脳内の信号処理と伝達に関係する脳科学の基礎を一般の人にもわかるようにできるだけ、
数式と回路を用いないで説明します
しかし、脳内の信号処理と伝達を科学的に分析するためには、
信号処理や伝達をできるだけ数式や回路によって記述する必要がある
なので数式や回路による記述は、別の記事で解説します
数式と回路を用いないで説明します
しかし、脳内の信号処理と伝達を科学的に分析するためには、
信号処理や伝達をできるだけ数式や回路によって記述する必要がある
なので数式や回路による記述は、別の記事で解説します
<< 神経細胞(ニューロン)と神経系について>>
1、神経細胞(ニューロン)
(1)神経細胞(ニューロン)は、脳内の情報処理と信号の伝達を担う素子である
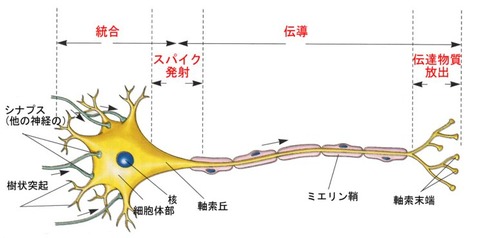
(2)神経細胞(ニューロン)の構造
①細胞体(soma)
細胞核(DNA)などが含まれている部分で、ニューロンの本体といえる部分。
②樹状突起(dendrite)
細胞体から伸びだした多数の枝のような部分で、ニューロンの入力端子にあたるところ。
他の細胞からの入力を受ける
③軸索(axon)
細胞体から伸びだし、一般に太くて枝わかれしない、
他の細胞に出力するニューロンの出力端子にあたる部分。
④シナプス(synapse)
ニューロン間で20nm程度の間隙はあるがほとんどつながっている結合部分
ニューロンとニューロンをつなげる役割をする。
化学物質を放出する側のシナプスを前シナプス、
化学物質を受け取る側のシナプスを後シナプスという
前シナプスは、軸索終末にあり、



(3)一つの化学信号によってシナプスに発生する電気信号
一つの化学信号によってシナプスに発生する電位変化の電気信号は、
下記の図などから推測で以下のものである






1、神経細胞(ニューロン)
(1)神経細胞(ニューロン)は、脳内の情報処理と信号の伝達を担う素子である
(2)神経細胞(ニューロン)の構造
①細胞体(soma)
細胞核(DNA)などが含まれている部分で、ニューロンの本体といえる部分。
②樹状突起(dendrite)
細胞体から伸びだした多数の枝のような部分で、ニューロンの入力端子にあたるところ。
他の細胞からの入力を受ける
③軸索(axon)
細胞体から伸びだし、一般に太くて枝わかれしない、
他の細胞に出力するニューロンの出力端子にあたる部分。
④シナプス(synapse)
ニューロン間で20nm程度の間隙はあるがほとんどつながっている結合部分
ニューロンとニューロンをつなげる役割をする。
化学物質を放出する側のシナプスを前シナプス、
化学物質を受け取る側のシナプスを後シナプスという
前シナプスは、軸索終末にあり、
後シナプスは、樹状突起付近にある


*** 参考資料 ***
脳内の情報処理機能を担う素子は神経細胞(ニューロン)である。
神経細胞同士の信号の伝達は、電気信号(活動電位)によって行われる。
活動電位が出なければ他の神経細胞に情報を伝達することができない。
活動電位が出てはじめて信号は神経線維を伝わり次の神経細胞へと伝達される。
脳内の記憶情報の処理も神経細胞の活動電位によって担われている。
一方、活動電位を出すか出さないかは、
シナプスに発生する信号の合成の結果が活動電位発生
の閾値に達するかどうかにかかっている。
http://www.pri.kyoto-u.ac.jp/brain/brain/45/index-45.html
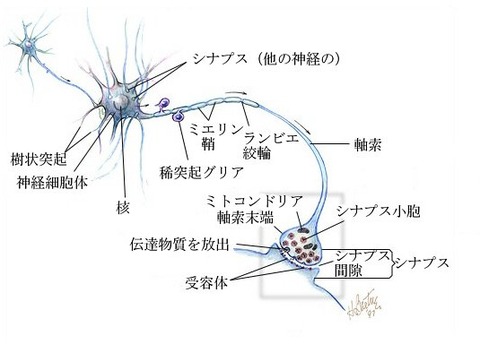
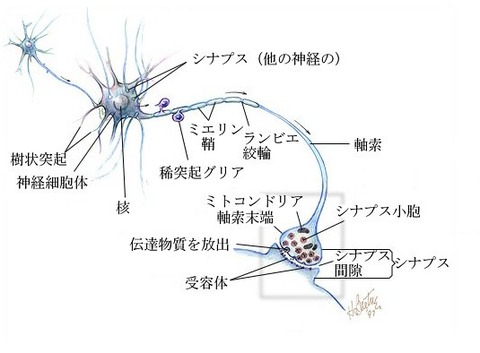
神経細胞(しんけいさいぼう、ニューロン、neuron)は、神経系を構成する細胞で、
その機能は情報処理と情報伝達に特化しており、動物に特有である。
神経細胞は主に3つの部分に区分けされ、細胞核のある細胞体、
他の細胞からの入力を受ける樹状突起、他の細胞に出力する軸索に分けられる。
前の細胞の軸索終末と後ろの細胞の樹状突起の間の情報を伝達する部分には、
微小な間隙を持つシナプスと呼ばれる化学物質による伝達構造が形成されている。
http://ja.wikipedia.org/wiki/%E3%83%8B%E3%83%A5%E3%83%BC%E3%83%AD%E3%83%B3
神経の基本単位はニューロン(神経単位, neuron)と呼ばれ、
細胞体 (soma)、多数の樹状突起 (dendrite)、軸索 (axon)、
神経終末 (nerve terminal)からなる。
軸索は情報を伝えるケーブルで神経線維とも呼ばれ、
電気的絶縁性の高い髄鞘の有無でそれぞれ有髄神経と無髄神経に分類される。
髄鞘は1-2mm間隔で細胞膜が露出して、この露出部をランビエ絞輪と呼ぶ。
http://sugp.int-univ.com/Material/Medicine/cai/text/subject01/no1/html/section2.html
ニューロンはそれぞれの部域で機能的な役割の分担をしている。
樹状突起部は、他からの情報を受け取る場所である。
樹状突起(と細胞体)には多数のシナプスが形成されている。
情報はここから入力される。細胞体部はおもに樹状突起で受けた入力を統合する場所となる。
http://www.tmd.ac.jp/artsci/biol/textlife/neuron.htm#top
神経細胞(ニューロン)の構造
1、細胞体(soma)
核などが含まれている部分で、ニューロンの本体といえる部分。
2、樹状突起(dendrite)
細胞体から伸びだした多数の枝のような部分で、ニューロンの入力端子にあたるところ。
3、軸索(axon)
細胞体から伸びだし、一般に太くて枝わかれしない、ニューロンの出力端子にあたる部分。
4、シナプス(synapse)
他のニューロンをつなげる役割をする。樹状突起はシナプスを通して、
他のニューロンから入力信号を受け取る。
また、シナプスの伝達効率はそれぞれ異なっている。
http://mars.elcom.nitech.ac.jp/java-cai/neuro/neuro1.html
化学シナプスの基本的構造は、神経細胞の軸索の先端が
他の細胞(神経細胞の樹状突起や筋線維)と
20nm程度の隙間(シナプス間隙)を空けて、
シナプス接着分子によって細胞接着している状態である。
http://ja.wikipedia.org/wiki/%E3%82%B7%E3%83%8A%E3%83%97%E3%82%B9
2、神経系
神経系(しんけいけい)とは、動物のうち、多くのグループに見られる器官系のひとつ。
主に神経細胞の働きによって情報の伝達と処理を行う一連の器官のこと。
神経系の構造と機能について研究する学問は、脳科学または神経科学と呼ばれる。
神経系 - 中枢神経 - 脳
l l
l └ 脊髄
l
└ 末梢神経 - 体性神経 - 感覚神経
l l
l └ 運動神経
l
└ 自律神経 - 副交感神経
l
└ 交感神経
末梢神経 - 脳神経 = 脳から直接出ている神経
l
└ 脊髄神経 = 脊髄から出ている神経
脳神経 = 脳から直接出ている神経 = 視覚情報、聴覚情報、嗅覚情報などの神経
脊髄神経 = 脊髄から出ている神経 = 運動神経など
神経系 = 全ての信号の伝達網と信号処理を担うネットワーク
信号は全て電気信号と化学信号によって伝達、処理される
中枢神経 = 神経細胞(ニューロン)の集合体
脊髄 = 脳からの命令を各部に伝え、各部から得た情報を脳へ伝える
感覚神経 = 感覚情報を脳との間で伝達する
運動神経 = 運動情報を脳との間で伝達する
自律神経 = 心拍、呼吸、分泌の調節など、内部環境の調整を自動的に行う
脳 = 神経細胞(ニューロン) + グリア細胞 で構成されている
神経細胞は大脳で数百億個、小脳で千億個ある
グリア細胞は脳内に神経細胞の10倍以上ある
神経細胞(ニューロン)= 信号の伝達、処理を担当
グリア細胞
= 神経細胞に寄り添うようにあり、
神経細胞に栄養分を供給したり、損傷した神経細胞を修復したりするなど
神経細胞の働きを支え、メンテナンスの役割を担っている
*** 参考資料 ***
神経系(しんけいけい)とは、動物のうち、多くのグループに見られる器官系のひとつ。
主に神経細胞の働きによって情報の伝達と処理を行う一連の器官のこと。
神経系の構造と機能について研究する学問は、脳科学または神経科学と呼ばれる。
http://ja.wikipedia.org/wiki/%E7%A5%9E%E7%B5%8C%E7%B3%BB
参考文献:「脳のしくみがわかる本」 寺沢宏次 監修
<<膜電位、イオンポンプ、イオンチャネル、イオン電流、平衡電位について>>
1、静止膜電位
電荷を持つイオンの分布が細胞内外で異なるため、
神経細胞の内と外の間では、電位差が存在している。
活性化してない静止状態の細胞の電位差は静止膜電位(または静止電位)といい、
通常細胞外と比べ細胞内がマイナスとなっている。
この状態を、膜は分極しているという。
神経細胞ではこの静止電位は、およそ-70mVである(細胞外の電位を0としている)。
***参考資料***
膜電位
膜電位(まくでんい; membrane potential)は細胞の内外に存在する電位の差のこと。
すべての細胞は細胞膜をはさんで細胞の中と外とでイオンの組成が異なっており、
この電荷を持つイオンの分布の差が、電位の差をもたらす。
通常、細胞内は細胞外に対して負(陰性)の電位にある。
細胞内外に濃度差を作られたイオンは電荷を持っているので、内外のイオンバランスの差は、
内外の電気的ポテンシャルの差をもたらす。つまり、イオンの分布差そのものが、
細胞内外に電位の差をもたらすということである。
この、イオン分布の差による細胞内外の電位差を、膜電位と呼ぶのである。
仮に膜外に100個の1価の陽イオンがあり、
膜内に40個の1価の陽イオンがあるという状況を想定する。
この場合、膜外は膜内に対して、イオン60個分のプラスの電位差を持っているといえる
(逆に、膜内は膜外に比べ、イオン60個分マイナスの電位差があるといえる)。
このように、膜電位とは膜内外の陰陽両イオンの電荷の総和で決定される。
現実には膜内外に存在するイオンは一種類ではなく、
またイオン種によって価数も違うため、計算は容易ではない。
また、電荷バランスが崩れた領域は、膜の近傍の2~3nm(デバイ長)のところのみである。
したがって、大部分の電荷は膜表面付近に集中する。
http://ja.wikipedia.org/wiki/%E8%86%9C%E9%9B%BB%E4%BD%8D
活動電位について知るためには、
まず静止膜電位(静止電位ともいう)についておおまかに知る必要がある。
すべての細胞の膜内外に存在する電位差は通常、細胞外と比べ細胞内がマイナスである。
この状態を、膜は分極しているという。
活動していない状態にある膜の電位差は静止電位と呼ばれ、
神経細胞ではおよそ-70mVである(細胞外の電位を0としている)。
この電位差が生ずる要因はいくつかあるが、
もっとも重要なものは膜間における、イオン輸送と選択的なイオン透過性である。
活動電位の大まかな流れは次のようになっている。
静止電位
静止電位において、いくつかのK+漏洩チャネルは開いている一方、
電位依存性Na+チャネルは閉じている。正味の電流は流れていないが、
膜間を移動している主なイオンはK+であり、
その結果静止電位はK+平衡電位に比較近い値をとる。
http://ja.wikipedia.org/wiki/%E6%B4%BB%E5%8B%95%E9%9B%BB%E4%BD%8D
2、イオンポンプ
神経細胞には、ナトリウム-カリウムポンプ
(Na+-K+ ATPアーゼ; ナトリウムポンプとも)と呼ばれる
ナトリウム(Na+)を細胞外に排出し、カリウム(K+)を
細胞内に入れるという作業をする部分がある
これはATPの加水分解によるエネルギーを利用して
3個のナトリウムイオン(Na+)を細胞外に汲み出すと共に、
2個のカリウムイオン(K+)を細胞内に汲み込むタンパクである。
したがって、3個のプラス電荷が細胞外に排出され、2個のプラス電荷が細胞内に流入するので
細胞内の電荷 = +2(2個のカリウムイオン:K+)-3(3個のナトリウムイオン:Na+)
= -1
となりどんどん細胞内のプラス電荷が少なくなり、
細胞内の電位は細胞外の電位と比較してマイナスになっていく
最終的に、細胞内にはカリウムイオンが多くなり、細胞外にはナトリウムイオンが多いという
平衡状態が生まれ細胞内外で電荷分布が異なり
神経細胞の内と外の間では、電位差が存在することになる
これが神経細胞において静止膜電位が生じる原因であり、
神経細胞に電気信号を生じさせるエネルギー源である
***参考資料***
イオンポンプ
膜電位に関わるイオンポンプとして、もっとも有名かつ研究がなされたものとして、
ナトリウム-カリウムポンプ(Na+-K+ ATPアーゼ; ナトリウムポンプとも)が挙げられる(右図)
。 これはATPの加水分解によるエネルギーを利用して
3個のナトリウムイオン(Na+)を細胞外に汲み出すと共に、
2個のカリウムイオン(K+)を細胞内に汲み込むタンパクである。
このタンパクが働いているおかげで、細胞内はナトリウムイオンが少なく、
カリウムイオンが多いという条件を維持できるのである。
そのほかにもカルシウムイオン(Ca2+)や水素イオン(H+)を輸送するポンプなども存在し、
成分としては小さいものの、膜電位に貢献している。
http://ja.wikipedia.org/wiki/%E8%86%9C%E9%9B%BB%E4%BD%8D
3、イオンチャネル
イオンチャネル (ion channel) とは、細胞の生体膜(細胞膜や内膜など)にある
膜貫通タンパク質で、特定のイオンを透過させる働きを有するものの総称である。
イオンの選択性はチャネルによってまちまちであり、
一種類のイオンのみ選択的に透過させるチャネルもあれば、
多くの陽イオンを通してしまうチャネルも存在する。
神経細胞には以下の種類のイオンだけを通すイオンチャネルがある
(1)Na+チャネル(ナトリウムチャネル;sodium channel)、
(2)Ca2+チャネル(カルシウムチャネル; calcium channel)、
(3)K+チャネル(カリウムチャネル;potassium channel)、
(4)塩素イオン(Cl-)チャネル(chloride channel)
(5)K+とCl-チャネル(K+とCl-の2つのイオンだけを通すチャネル)
(6)Na+とK+チャネル(Na+とK+の2つのイオンだけを通すチャネル)
***参考資料***
膜に存在するタンパク質で、刺激に応じて開閉しイオンが通過する小孔を形成する。
透過するイオンの選択性によりNa+チャネル(ナトリウムチャネル;sodium channel)、
Ca2+チャネル(カルシウムチャネル; calcium channel)、
K+チャネル(カリウムチャネル;potassium channel)、
塩素イオンチャネル(chloride channel)と名づけられている。
イオンは電気化学的勾配に従って移動し、
この際、ATPなどのエネルギーは、必要としない(受動輸送)。
膜内外の電位差を感知して開閉する
電位依存型チャネルと内因性物質を感知して開閉する
リガンド依存性チャネルがある。
リガンド依存性チャネルはイオンチャネル内蔵型受容体とも呼ばれる。
電位依存性Na+チャネルは神経や筋肉の細胞膜に存在し、
細胞膜の脱分極により活性化される。
細胞内へのNa+イオンの透過性を高め、神経や筋肉に活動電位を発生させる。
フグ毒のテトロドトキシンは、このチャネルを塞ぐことにより、
運動神経・骨格筋の興奮を抑え、呼吸麻痺をもたらす。
ニコチン型アセチルコリン受容体はNa+イオンを透過させる
リガンド依存性Na+チャネルである。
電位依存性Ca2+チャネルには、L、T、N、P/Q、R型などがあり、それぞれ発現部位が異なる。
膜の脱分極により活性化しCa2+イオンを透過させる。
L型Ca2+チャネルは骨格筋・心筋・血管・脳など、多くの興奮性細胞に存在して、
細胞外から細胞内に、Ca2+(カルシウムイオン)を流入させる。
Caブロッカー(Ca2+拮抗薬;カルシウム拮抗薬)は、
L型Ca2+チャネルのα1サブユニットに結合し、
血管平滑筋へのCa2+流入を阻害し、降圧効果を現わす。
N型Ca2+チャネルは神経に存在する。
K+チャネルは、膜電位調節に大きく関わっている。
電位依存性K+チャネルは中枢及び末梢神経・心筋細胞など
にあり細胞膜の脱分極によって開口する。
活動電位の持続時間、発現間隔を規定する。
電位依存性K+チャネル阻害薬は心臓のクラスIII群抗不整脈薬が代表的である。
内向き整流性K+チャネルにはATP感受性K+チャネル、Gタンパク質制御K+チャネルがある。
膵β細胞では血糖値上昇に伴い細胞内ATPが上昇し、ATP感受性K+チャネルが閉じ、
細胞膜が脱分極する。その結果インスリンの分泌が促進される。
http://www.pharm.or.jp/dictionary/wiki.cgi?%E3%82%A4%E3%82%AA%E3%83%B3%E3%83%81%E3%83%A3%E3%83%8D%E3%83%AB
4、イオンチャネルの開閉制御方法
イオンチャネルが開いたり閉じたりするのを制御する方法には以下の種類がある
(1)電位依存性 (voltage-dependent channel/voltage-gated channel)
=膜電位の変化に応じて開くもの。
時定数の異なる複数のゲートを持ち、
膜電位変化時に時間に依存した特定の開閉を行うチャネルも多い。
神経細胞のシナプス内のイオン電荷によって発生するシナプス電位があるが、
このシナプス電位に応じて活動電位を発生させるイオンチャネルは、
この電位依存性チャネルで、細胞内にイオンが流入してプラス電位を発生させるNa+チャネルと
細胞外にイオンが流出してマイナス電位を発生させるK+チャネルがある
(2)リガンド依存性(ligand-gated channel):特異的に結合する分子によるもの。
この場合イオンチャネル自体が受容体となっている。
受容体の側から見れば、イオンチャネル共役型受容体とも呼べる。
たとえば、AMPA型グルタミン酸受容体、NMDA型グルタミン酸受容体など。
脳内において神経細胞のシナプス間を神経伝達物質が伝導して、神経伝達物質によって
シナプス内に電位を発生させているイオンチャネルは、
このリガンド依存性チャネルであり、
細胞内にプラス電位を発生させるNa+とK+を通すチャネルと
細胞内にプラス電位を発生させるのを抑制するCl-チャネルがある
(3)リン酸化依存性:他タンパクからのリン酸化シグナルによるもの
(4)力学的変形依存性:チャネル分子に機械的変形が関わると開くもの。
聴覚器官における内耳の有毛細胞のチャネルが、力学的変形依存性チャネルである
音波の機械的振動によって有毛細胞の毛が振動し、
その毛の振動に応じてチャネルが開閉し、細胞内にイオンを流入させる
(5)温度依存性:温度によるもの。
(6)漏洩チャネル:通常開いており、少しずつ特定のイオンを漏らすように流すもの。
シナプスには、K+とCl-のイオンが通れる漏洩チャネルがあり
常にK+イオンとCl-イオンが細胞内外を移動している
(7)光依存性:光量によりチャネルを開閉する
網膜の細胞に存在している
網膜の視細胞は、光量によってチャネルは確かに開閉するが、
その開閉過程は少し複雑で、以下のようにして開閉する
暗所においてはcGMPがイオンチャネルに結合し、イオンチャネルは
Na、Caイオンに対して開かれている。
そのため内部のNa濃度は高く、桿体細胞は脱分極の状態にある。
光が桿体にあたると光感受性のあるレチナールが光子を吸収し、
シス型からall-trans型に異性体変異を起こす。
そのときビタミンAは伸張するため、ロドプシンの構造が変化し、
それが周辺にあるトランスデューシンG蛋白を刺激する。
αβγという3部品からなっているG蛋白からαサブユニットが解離する。
解離したαはフォスフォジエステラーゼ という酵素を活性化し、cGMPを分解する。
その結果イオンチャネルが閉じ、過分極を起こす。
http://www.ccn.yamanashi.ac.jp/~k-sakai/visual.html
***参考資料***
制御様式
イオンチャネルの開閉の制御様式には、いくつかある。
電位依存性 (voltage-dependent channel/voltage-gated channel)
:膜電位の変化に応じて開くもの。
時定数の異なる複数のゲートを持ち、
膜電位変化時に時間に依存した特定の開閉を行うチャネルも多い。
リガンド依存性(ligand-gated channel):特異的に結合する分子によるもの。
この場合イオンチャネル自体が受容体となっている。受容体の側から見れば、
イオンチャネル共役型受容体とも呼べる。
たとえば、AMPA型グルタミン酸受容体、NMDA型グルタミン酸受容体など。
リン酸化依存性:他タンパクからのリン酸化シグナルによるもの
力学的変形依存性:チャネル分子に機械的変形が関わると開くもの。
内耳の有毛細胞などが有名。
温度依存性:温度によるもの。
漏洩チャネル:通常開いており、少しずつ特定のイオンを漏らすように流すもの。
http://ja.wikipedia.org/wiki/%E3%82%A4%E3%82%AA%E3%83%B3%E3%83%81%E3%83%A3%E3%83%8D%E3%83%AB
活動電位からシナプス電位へ
シナプスにおいて、活動電位からシナプス電位に変換されます(図5)。
活動電位は、神経細胞の軸索突起の根元で発生し、軸索をつたわってシナプス前部に達します。
シナプス前部には膜電位感受性カルシウムチャネルがありますので、
これが活動電位によって膜電位が上昇したことを感知して、活性化し、
つまりチャネルが開いてカルシウムが細胞内にはいります。
このカルシウムによって、シナプス前部にあったシナプス小胞が細胞膜に融合し、
中に入っていた神経伝達物質をシナプス間隙に放出します。
興奮性の神経伝達を司っているグルタミン酸を例にあげてみますと、
グルタミン酸がシナプス後膜にあるその受容体、つまりグルタミン酸受容体に結合します。
グルタミン酸受容体は4つの膜蛋白からなり、それらのまん中に穴(チャネル)を形成していて、
受容体にグルタミン酸が結合するとタンパク質の構造が変化して、短時間穴が開きます。
この穴はカチオン(陽イオン;ナトリウム、カリウム、カルシウムなど)をとおす性質があるため、
細胞外には内側に比べて高濃度のナトリウムイオンがあり、
それがチャネルをとおって細胞内に流入します。ナトリウムイオンはプラスですから、
その結果、細胞内のマイナスの程度が浅くなる、つまり脱分極します。
脱分極するにつれ、カリウムイオンの流出もあるために、
活動電位のように膜電位がプラスまで上昇することはありません。
このような神経伝達物質によって引き起されたシナプス後部の膜電位をシナプス電位といいます。
グルタミン酸 (Glu) の場合、シナプス電位は脱分極性なので興奮性シナプス後電位といいます。
一方、GABA (ギャバ) やグリシンは抑制性です。
抑制性ニューロンからGABAが分泌されると、GABA受容体チャネルが活性化し、
塩素イオンを選択的に通す穴がひらき、細胞外に多い塩素イオンが細胞内に流入します。
塩素イオンはマイナスなので、その結果膜電位はよりマイナスの方向に深くなり、
つまり過分極になり、これを抑制性シナプス後電位といいます。
これによって、活動電位が出にくくなります(図5)。
麻酔薬、睡眠薬、抗不安薬などはこのGABA受容体の働きを促進させ、
グルタミン酸による興奮性の神経活動を抑制させるのです。
http://www.tmin.ac.jp/medical/17/synapse2.html
AMPA型グルタミン酸受容体
電流-電圧特性
GluR2を持つ受容体の他の相違点としては、比較的オームの法則に従った、
線形の電流-電圧特性を持つことがあげられる。
他のサブユニットのみで構成されるAMPA受容体は、
膜電位が負の状態ではオームの法則に従うが、正の状態ではほとんど電流を流さない、
内向きの整流性を持つことが知られている(右図参照)。
これは、膜電位が正のとき、GluR1,3,4の各サブユニットは
細胞内ポリアミンによる阻害を受けているためである。
http://ja.wikipedia.org/wiki/AMPA%E5%9E%8B%E3%82%B0%E3%83%AB%E3%82%BF%E3%83%9F%E3%83%B3%E9%85%B8%E5%8F%97%E5%AE%B9%E4%BD%93
NMDA型グルタミン酸受容体
NMDA受容体は通常不活性な性質を持つ。
これは、細胞外からのマグネシウムイオンがこの受容体の活動を阻害しているためである
(マグネシウム・ブロック)。ただし、これは膜電位が大きく負の時に限られている。
そのため、膜電位が正であるか、-10~-20 mV 程度の範囲においては
マグネシウムイオン阻害がかからず、
結果として受容体の電気生理学的特性はオームの法則に従う。
膜電位が大きく負になるとマグネシウムイオンの阻害がかかり始め、
-60~-70 mV 程度の静止膜電位に相当する膜電位では、ほとんど電流を流さない。
これらのことから、NMDA型受容体は外向き整流性を持つ、と称される。
また、刺激に応じて流す電流は、AMPA受容体に比べて遅く、持続的である。
http://ja.wikipedia.org/wiki/NMDA%E5%9E%8B%E3%82%B0%E3%83%AB%E3%82%BF%E3%83%9F%E3%83%B3%E9%85%B8%E5%8F%97%E5%AE%B9%E4%BD%93
5、シナプスのイオンチャネル
(1)活動電位を発生させるイオンチャネル=電位依存性チャネル
シナプス電位に応じてチャネルを開閉しており以下の2種類ある
①細胞内にイオンが流入してプラス電位を発生させるNa+チャネル
②細胞外にイオンが流出してマイナス電位を発生させるK+チャネル
(2)通常の神経細胞のシナプス電位を発生させるイオンチャネル
=リガンド依存性チャネルと漏洩チャネル
(2-1)リガンド依存性チャネル
脳内において神経細胞のシナプス間を神経伝達物質が伝導して、神経伝達物質によって
シナプス内に電位を発生させているイオンチャネルは、
リガンド依存性チャネルであり、以下の2種類がある
①プラス電位を発生させるNa+とK+チャネル
これは1つのチャネルでNa+とK+の2つを通すということである
②Cl-チャネル
Cl-チャネルは、静止膜電位と同じ電位を発生させる
つまり、プラス電位を発生させるのを抑制するような働きがある
(2-2)漏洩チャネル
シナプスには、K+とCl-のイオンが通れる漏洩チャネルがあり
常にK+イオンとCl-イオンが細胞内外を、
イオン濃度と膜電位に応じて自由に移動している
この漏洩チャネルが-70mvの静止膜電位を発生させている原因となっている
つまり、イオンポンプが
3個のナトリウムイオン(Na+)を細胞外に汲み出すと共に、
細胞内に2個のK+イオンを流入させている一方で、
K+の漏洩チャネルによりK+イオンが細胞外へと流出させている
以下にその関係を単純化させた式を示す
細胞内の電荷
=+2(イオンポンプによる2個のK+流入)
-3(イオンポンプによる3個のNa+流出)
-1(K+の漏洩チャネルによりK+流出)
=-2
つまりイオンポンプだけなら、-70mvよりも高い電位であったろうが
K+の漏洩チャネルによりK+イオンが流出して-70mvになっている
***参考資料***
漏洩チャネルの貢献
神経細胞の典型的な軸索において、静止膜電位は負であり、おおよそ-70mV程度である。
このことは、細胞外に陽イオンが比較的多い(もしくは細胞内に陰イオンが比較的多い)
ことを示唆している。実際には前者が正しい。
前述したNa+-K+交換イオンポンプは3個のナトリウムイオンと2個のカリウムイオンを
交換しているだけなので膜電位の変化にはそれほど大きく寄与しないが、
外にくみ出されたナトリウムイオンが細胞内に入り込むための
ナトリウムチャネルは通常不活性化されており、
開いていないのに対し、カリウムが細胞外に流出するカリウムチャネルの中には、
通常開きっぱなしのものが存在する。つまり、カリウムイオンは汲み入れても汲み入れても、
ある程度は細胞外に漏れ出て行ってしまうのである。
これが、静止膜電位が負になってしまう主な原因である。
このカリウムイオンを漏れ出させてしまうチャネルを、カリウム漏洩チャネルと呼ぶ。
http://ja.wikipedia.org/wiki/%E8%86%9C%E9%9B%BB%E4%BD%8D
塩化物イオン(Cl-)の移動は細胞膜を通してかなり自由度が高いため、
この膜電位による移動にほとんど依存している。
http://ja.wikipedia.org/wiki/%E8%86%9C%E9%9B%BB%E4%BD%8D
6、イオンチャネルを流れるイオン電流
イオンを流す方向は、チャネルによって決める事はできない。
なぜなら、イオンチャネルはイオンを受動的に輸送するからである。
イオンチャネルが開くと、イオンは以下の電流が流れる
(1)膜内外のイオンの濃度差により濃度を平均化しようとして流れる拡散電流
(2)膜電位差による電圧電流
***参考資料***
イオンの選択性はチャネルによってまちまちであり、
一種類のイオンのみ選択的に透過させるチャネルもあれば、
多くの陽イオンを通してしまうチャネルも存在する。
しかし、イオンを流す方向は、チャネルによって決める事はできない。
なぜなら、イオンチャネルはイオンを受動的に輸送するからである。
イオンチャネルが開くと、イオンは電気化学的勾配の高い方から低い方へ流れる。
つまり、イオンチャネルがいくら開いても、膜電位が平衡電位と等しい場合、
すなわち、膜内外でのイオン濃度差による拡散の効果と、
膜電位によるクーロンポテンシャルの効果が
釣り合っているとイオン電流は流れない。
http://ja.wikipedia.org/wiki/%E3%82%A4%E3%82%AA%E3%83%B3%E3%83%81%E3%83%A3%E3%83%8D%E3%83%AB
イオンチャネルによるイオンの移動
次に、イオンポンプなどの活動により一旦イオン分布の差が生まれると、
今度はその濃度差を利用した受動輸送が可能になる。
この受動輸送は、イオンチャネルと呼ばれるタンパク質によってなされる。
イオンチャネルは、イオンポンプ等によって濃度差が作られたイオンを、
イオン濃度の高いほうから低いほうへ拡散させる、イオンの通り道である。
よって、方向に選択性はなく、
膜電位がない場合は常にイオンの濃度勾配に従った輸送である。
ただし、イオン濃度の低い方から高い方への移動が
全くないわけではないことに注意すべきである。
イオンがチャネルを通過するかどうかはそのイオンがブラウン運動によって
チャネル分子に衝突するかどうかに依存しており、イオン濃度の高い側では、
イオンのチャネルへの衝突が、低いほうに比べて圧倒的に起きやすいため、
全体としては高い方から低い方への流れが生じるわけである。
イオンチャネルの多くは通常不活性型であり、
何らかの刺激(膜電位の変化・リガンドの結合・リン酸化・機械刺激など)に
応じて開閉する。
そのため、定常状態の細胞において、
働いているイオンチャネルは少ないと言える。
ただし、漏洩チャネルと呼ばれるタイプのイオンチャネルは常に開いており、
静止膜電位に貢献する(後述)。
膜電位そのものによるイオンの移動
膜内外の電位差そのものも、イオンの移動に影響を及ぼす。
たとえば、静止状態の膜電位は細胞内が細胞外に比べて
負であることはすでに述べたが、
負の細胞内に向けて陽イオンは入りやすく、陰イオンは入りにくい。
逆に、正の細胞外に向けて陰イオンは出て行きやすく、陽イオンは出て行きにくい。
これは、単純に細胞外の正電荷を持つ環境が、陽イオンを反発させようとするからである。
現実に、塩化物イオン(Cl-)の移動は細胞膜を通してかなり自由度が高いため、
この膜電位による移動にほとんど依存している。
http://ja.wikipedia.org/wiki/%E8%86%9C%E9%9B%BB%E4%BD%8D
7、平衡電位と拡散電位
(1)平衡電位と拡散電位の説明
膜を通して流れる電流 = 拡散電流+電圧電流 = 0
となっている状態を平衡状態という
つまり平衡状態とは、
ある量Aと別のある量Bが一致して平衡しているときの状態をいう
この平衡状態では、拡散電流を阻止するように電圧電流が逆に流れているので
見かけ上は正味の電流は流れていない
この拡散電流と平衡する電圧電流の膜電位を、平衡電位といい
各イオンごとによって平衡電位が存在する
この平衡電位は、拡散電流を電圧電流と考えたときの電圧に相当するので、
拡散電位ともいう
ただし拡散電位というときは、
拡散電流(拡散電位電流) = -電圧電流(平衡電位電流)
なので平衡電位の逆の符号の電位となる
電気回路理論で、拡散電流を取り扱うときは
この平衡電位(拡散電位)の電圧(一般には化学電池と呼ばれている)
によって電流が流れるとして取り扱う
つまり、各イオンごとの平衡電位(拡散電位)が
拡散電流を流す電圧として考えることができるということである
(2)平衡電位の求め方(ネルンストの式)
各イオンごとの平衡電位の求め方は、以下のネルンストの式で求められる
脳内の情報処理機能を担う素子は神経細胞(ニューロン)である。
神経細胞同士の信号の伝達は、電気信号(活動電位)によって行われる。
活動電位が出なければ他の神経細胞に情報を伝達することができない。
活動電位が出てはじめて信号は神経線維を伝わり次の神経細胞へと伝達される。
脳内の記憶情報の処理も神経細胞の活動電位によって担われている。
一方、活動電位を出すか出さないかは、
シナプスに発生する信号の合成の結果が活動電位発生
の閾値に達するかどうかにかかっている。
http://www.pri.kyoto-u.ac.jp/brain/brain/45/index-45.html
神経細胞(しんけいさいぼう、ニューロン、neuron)は、神経系を構成する細胞で、
その機能は情報処理と情報伝達に特化しており、動物に特有である。
神経細胞は主に3つの部分に区分けされ、細胞核のある細胞体、
他の細胞からの入力を受ける樹状突起、他の細胞に出力する軸索に分けられる。
前の細胞の軸索終末と後ろの細胞の樹状突起の間の情報を伝達する部分には、
微小な間隙を持つシナプスと呼ばれる化学物質による伝達構造が形成されている。
http://ja.wikipedia.org/wiki/%E3%83%8B%E3%83%A5%E3%83%BC%E3%83%AD%E3%83%B3
神経の基本単位はニューロン(神経単位, neuron)と呼ばれ、
細胞体 (soma)、多数の樹状突起 (dendrite)、軸索 (axon)、
神経終末 (nerve terminal)からなる。
軸索は情報を伝えるケーブルで神経線維とも呼ばれ、
電気的絶縁性の高い髄鞘の有無でそれぞれ有髄神経と無髄神経に分類される。
髄鞘は1-2mm間隔で細胞膜が露出して、この露出部をランビエ絞輪と呼ぶ。
http://sugp.int-univ.com/Material/Medicine/cai/text/subject01/no1/html/section2.html
ニューロンはそれぞれの部域で機能的な役割の分担をしている。
樹状突起部は、他からの情報を受け取る場所である。
樹状突起(と細胞体)には多数のシナプスが形成されている。
情報はここから入力される。細胞体部はおもに樹状突起で受けた入力を統合する場所となる。
http://www.tmd.ac.jp/artsci/biol/textlife/neuron.htm#top
神経細胞(ニューロン)の構造
1、細胞体(soma)
核などが含まれている部分で、ニューロンの本体といえる部分。
2、樹状突起(dendrite)
細胞体から伸びだした多数の枝のような部分で、ニューロンの入力端子にあたるところ。
3、軸索(axon)
細胞体から伸びだし、一般に太くて枝わかれしない、ニューロンの出力端子にあたる部分。
4、シナプス(synapse)
他のニューロンをつなげる役割をする。樹状突起はシナプスを通して、
他のニューロンから入力信号を受け取る。
また、シナプスの伝達効率はそれぞれ異なっている。
http://mars.elcom.nitech.ac.jp/java-cai/neuro/neuro1.html
化学シナプスの基本的構造は、神経細胞の軸索の先端が
他の細胞(神経細胞の樹状突起や筋線維)と
20nm程度の隙間(シナプス間隙)を空けて、
シナプス接着分子によって細胞接着している状態である。
http://ja.wikipedia.org/wiki/%E3%82%B7%E3%83%8A%E3%83%97%E3%82%B9
2、神経系
神経系(しんけいけい)とは、動物のうち、多くのグループに見られる器官系のひとつ。
主に神経細胞の働きによって情報の伝達と処理を行う一連の器官のこと。
神経系の構造と機能について研究する学問は、脳科学または神経科学と呼ばれる。
神経系 - 中枢神経 - 脳
l l
l └ 脊髄
l
└ 末梢神経 - 体性神経 - 感覚神経
l l
l └ 運動神経
l
└ 自律神経 - 副交感神経
l
└ 交感神経
末梢神経 - 脳神経 = 脳から直接出ている神経
l
└ 脊髄神経 = 脊髄から出ている神経
脳神経 = 脳から直接出ている神経 = 視覚情報、聴覚情報、嗅覚情報などの神経
脊髄神経 = 脊髄から出ている神経 = 運動神経など
神経系 = 全ての信号の伝達網と信号処理を担うネットワーク
信号は全て電気信号と化学信号によって伝達、処理される
中枢神経 = 神経細胞(ニューロン)の集合体
脊髄 = 脳からの命令を各部に伝え、各部から得た情報を脳へ伝える
感覚神経 = 感覚情報を脳との間で伝達する
運動神経 = 運動情報を脳との間で伝達する
自律神経 = 心拍、呼吸、分泌の調節など、内部環境の調整を自動的に行う
脳 = 神経細胞(ニューロン) + グリア細胞 で構成されている
神経細胞は大脳で数百億個、小脳で千億個ある
グリア細胞は脳内に神経細胞の10倍以上ある
神経細胞(ニューロン)= 信号の伝達、処理を担当
グリア細胞
= 神経細胞に寄り添うようにあり、
神経細胞に栄養分を供給したり、損傷した神経細胞を修復したりするなど
神経細胞の働きを支え、メンテナンスの役割を担っている
*** 参考資料 ***
神経系(しんけいけい)とは、動物のうち、多くのグループに見られる器官系のひとつ。
主に神経細胞の働きによって情報の伝達と処理を行う一連の器官のこと。
神経系の構造と機能について研究する学問は、脳科学または神経科学と呼ばれる。
http://ja.wikipedia.org/wiki/%E7%A5%9E%E7%B5%8C%E7%B3%BB
参考文献:「脳のしくみがわかる本」 寺沢宏次 監修
<<膜電位、イオンポンプ、イオンチャネル、イオン電流、平衡電位について>>
1、静止膜電位
電荷を持つイオンの分布が細胞内外で異なるため、
神経細胞の内と外の間では、電位差が存在している。
活性化してない静止状態の細胞の電位差は静止膜電位(または静止電位)といい、
通常細胞外と比べ細胞内がマイナスとなっている。
この状態を、膜は分極しているという。
神経細胞ではこの静止電位は、およそ-70mVである(細胞外の電位を0としている)。
***参考資料***
膜電位
膜電位(まくでんい; membrane potential)は細胞の内外に存在する電位の差のこと。
すべての細胞は細胞膜をはさんで細胞の中と外とでイオンの組成が異なっており、
この電荷を持つイオンの分布の差が、電位の差をもたらす。
通常、細胞内は細胞外に対して負(陰性)の電位にある。
細胞内外に濃度差を作られたイオンは電荷を持っているので、内外のイオンバランスの差は、
内外の電気的ポテンシャルの差をもたらす。つまり、イオンの分布差そのものが、
細胞内外に電位の差をもたらすということである。
この、イオン分布の差による細胞内外の電位差を、膜電位と呼ぶのである。
仮に膜外に100個の1価の陽イオンがあり、
膜内に40個の1価の陽イオンがあるという状況を想定する。
この場合、膜外は膜内に対して、イオン60個分のプラスの電位差を持っているといえる
(逆に、膜内は膜外に比べ、イオン60個分マイナスの電位差があるといえる)。
このように、膜電位とは膜内外の陰陽両イオンの電荷の総和で決定される。
現実には膜内外に存在するイオンは一種類ではなく、
またイオン種によって価数も違うため、計算は容易ではない。
また、電荷バランスが崩れた領域は、膜の近傍の2~3nm(デバイ長)のところのみである。
したがって、大部分の電荷は膜表面付近に集中する。
http://ja.wikipedia.org/wiki/%E8%86%9C%E9%9B%BB%E4%BD%8D
活動電位について知るためには、
まず静止膜電位(静止電位ともいう)についておおまかに知る必要がある。
すべての細胞の膜内外に存在する電位差は通常、細胞外と比べ細胞内がマイナスである。
この状態を、膜は分極しているという。
活動していない状態にある膜の電位差は静止電位と呼ばれ、
神経細胞ではおよそ-70mVである(細胞外の電位を0としている)。
この電位差が生ずる要因はいくつかあるが、
もっとも重要なものは膜間における、イオン輸送と選択的なイオン透過性である。
活動電位の大まかな流れは次のようになっている。
静止電位
静止電位において、いくつかのK+漏洩チャネルは開いている一方、
電位依存性Na+チャネルは閉じている。正味の電流は流れていないが、
膜間を移動している主なイオンはK+であり、
その結果静止電位はK+平衡電位に比較近い値をとる。
http://ja.wikipedia.org/wiki/%E6%B4%BB%E5%8B%95%E9%9B%BB%E4%BD%8D
2、イオンポンプ
神経細胞には、ナトリウム-カリウムポンプ
(Na+-K+ ATPアーゼ; ナトリウムポンプとも)と呼ばれる
ナトリウム(Na+)を細胞外に排出し、カリウム(K+)を
細胞内に入れるという作業をする部分がある
これはATPの加水分解によるエネルギーを利用して
3個のナトリウムイオン(Na+)を細胞外に汲み出すと共に、
2個のカリウムイオン(K+)を細胞内に汲み込むタンパクである。
したがって、3個のプラス電荷が細胞外に排出され、2個のプラス電荷が細胞内に流入するので
細胞内の電荷 = +2(2個のカリウムイオン:K+)-3(3個のナトリウムイオン:Na+)
= -1
となりどんどん細胞内のプラス電荷が少なくなり、
細胞内の電位は細胞外の電位と比較してマイナスになっていく
最終的に、細胞内にはカリウムイオンが多くなり、細胞外にはナトリウムイオンが多いという
平衡状態が生まれ細胞内外で電荷分布が異なり
神経細胞の内と外の間では、電位差が存在することになる
これが神経細胞において静止膜電位が生じる原因であり、
神経細胞に電気信号を生じさせるエネルギー源である
***参考資料***
イオンポンプ
膜電位に関わるイオンポンプとして、もっとも有名かつ研究がなされたものとして、
ナトリウム-カリウムポンプ(Na+-K+ ATPアーゼ; ナトリウムポンプとも)が挙げられる(右図)
。 これはATPの加水分解によるエネルギーを利用して
3個のナトリウムイオン(Na+)を細胞外に汲み出すと共に、
2個のカリウムイオン(K+)を細胞内に汲み込むタンパクである。
このタンパクが働いているおかげで、細胞内はナトリウムイオンが少なく、
カリウムイオンが多いという条件を維持できるのである。
そのほかにもカルシウムイオン(Ca2+)や水素イオン(H+)を輸送するポンプなども存在し、
成分としては小さいものの、膜電位に貢献している。
http://ja.wikipedia.org/wiki/%E8%86%9C%E9%9B%BB%E4%BD%8D
3、イオンチャネル
イオンチャネル (ion channel) とは、細胞の生体膜(細胞膜や内膜など)にある
膜貫通タンパク質で、特定のイオンを透過させる働きを有するものの総称である。
イオンの選択性はチャネルによってまちまちであり、
一種類のイオンのみ選択的に透過させるチャネルもあれば、
多くの陽イオンを通してしまうチャネルも存在する。
神経細胞には以下の種類のイオンだけを通すイオンチャネルがある
(1)Na+チャネル(ナトリウムチャネル;sodium channel)、
(2)Ca2+チャネル(カルシウムチャネル; calcium channel)、
(3)K+チャネル(カリウムチャネル;potassium channel)、
(4)塩素イオン(Cl-)チャネル(chloride channel)
(5)K+とCl-チャネル(K+とCl-の2つのイオンだけを通すチャネル)
(6)Na+とK+チャネル(Na+とK+の2つのイオンだけを通すチャネル)
***参考資料***
膜に存在するタンパク質で、刺激に応じて開閉しイオンが通過する小孔を形成する。
透過するイオンの選択性によりNa+チャネル(ナトリウムチャネル;sodium channel)、
Ca2+チャネル(カルシウムチャネル; calcium channel)、
K+チャネル(カリウムチャネル;potassium channel)、
塩素イオンチャネル(chloride channel)と名づけられている。
イオンは電気化学的勾配に従って移動し、
この際、ATPなどのエネルギーは、必要としない(受動輸送)。
膜内外の電位差を感知して開閉する
電位依存型チャネルと内因性物質を感知して開閉する
リガンド依存性チャネルがある。
リガンド依存性チャネルはイオンチャネル内蔵型受容体とも呼ばれる。
電位依存性Na+チャネルは神経や筋肉の細胞膜に存在し、
細胞膜の脱分極により活性化される。
細胞内へのNa+イオンの透過性を高め、神経や筋肉に活動電位を発生させる。
フグ毒のテトロドトキシンは、このチャネルを塞ぐことにより、
運動神経・骨格筋の興奮を抑え、呼吸麻痺をもたらす。
ニコチン型アセチルコリン受容体はNa+イオンを透過させる
リガンド依存性Na+チャネルである。
電位依存性Ca2+チャネルには、L、T、N、P/Q、R型などがあり、それぞれ発現部位が異なる。
膜の脱分極により活性化しCa2+イオンを透過させる。
L型Ca2+チャネルは骨格筋・心筋・血管・脳など、多くの興奮性細胞に存在して、
細胞外から細胞内に、Ca2+(カルシウムイオン)を流入させる。
Caブロッカー(Ca2+拮抗薬;カルシウム拮抗薬)は、
L型Ca2+チャネルのα1サブユニットに結合し、
血管平滑筋へのCa2+流入を阻害し、降圧効果を現わす。
N型Ca2+チャネルは神経に存在する。
K+チャネルは、膜電位調節に大きく関わっている。
電位依存性K+チャネルは中枢及び末梢神経・心筋細胞など
にあり細胞膜の脱分極によって開口する。
活動電位の持続時間、発現間隔を規定する。
電位依存性K+チャネル阻害薬は心臓のクラスIII群抗不整脈薬が代表的である。
内向き整流性K+チャネルにはATP感受性K+チャネル、Gタンパク質制御K+チャネルがある。
膵β細胞では血糖値上昇に伴い細胞内ATPが上昇し、ATP感受性K+チャネルが閉じ、
細胞膜が脱分極する。その結果インスリンの分泌が促進される。
http://www.pharm.or.jp/dictionary/wiki.cgi?%E3%82%A4%E3%82%AA%E3%83%B3%E3%83%81%E3%83%A3%E3%83%8D%E3%83%AB
4、イオンチャネルの開閉制御方法
イオンチャネルが開いたり閉じたりするのを制御する方法には以下の種類がある
(1)電位依存性 (voltage-dependent channel/voltage-gated channel)
=膜電位の変化に応じて開くもの。
時定数の異なる複数のゲートを持ち、
膜電位変化時に時間に依存した特定の開閉を行うチャネルも多い。
神経細胞のシナプス内のイオン電荷によって発生するシナプス電位があるが、
このシナプス電位に応じて活動電位を発生させるイオンチャネルは、
この電位依存性チャネルで、細胞内にイオンが流入してプラス電位を発生させるNa+チャネルと
細胞外にイオンが流出してマイナス電位を発生させるK+チャネルがある
(2)リガンド依存性(ligand-gated channel):特異的に結合する分子によるもの。
この場合イオンチャネル自体が受容体となっている。
受容体の側から見れば、イオンチャネル共役型受容体とも呼べる。
たとえば、AMPA型グルタミン酸受容体、NMDA型グルタミン酸受容体など。
脳内において神経細胞のシナプス間を神経伝達物質が伝導して、神経伝達物質によって
シナプス内に電位を発生させているイオンチャネルは、
このリガンド依存性チャネルであり、
細胞内にプラス電位を発生させるNa+とK+を通すチャネルと
細胞内にプラス電位を発生させるのを抑制するCl-チャネルがある
(3)リン酸化依存性:他タンパクからのリン酸化シグナルによるもの
(4)力学的変形依存性:チャネル分子に機械的変形が関わると開くもの。
聴覚器官における内耳の有毛細胞のチャネルが、力学的変形依存性チャネルである
音波の機械的振動によって有毛細胞の毛が振動し、
その毛の振動に応じてチャネルが開閉し、細胞内にイオンを流入させる
(5)温度依存性:温度によるもの。
(6)漏洩チャネル:通常開いており、少しずつ特定のイオンを漏らすように流すもの。
シナプスには、K+とCl-のイオンが通れる漏洩チャネルがあり
常にK+イオンとCl-イオンが細胞内外を移動している
(7)光依存性:光量によりチャネルを開閉する
網膜の細胞に存在している
網膜の視細胞は、光量によってチャネルは確かに開閉するが、
その開閉過程は少し複雑で、以下のようにして開閉する
暗所においてはcGMPがイオンチャネルに結合し、イオンチャネルは
Na、Caイオンに対して開かれている。
そのため内部のNa濃度は高く、桿体細胞は脱分極の状態にある。
光が桿体にあたると光感受性のあるレチナールが光子を吸収し、
シス型からall-trans型に異性体変異を起こす。
そのときビタミンAは伸張するため、ロドプシンの構造が変化し、
それが周辺にあるトランスデューシンG蛋白を刺激する。
αβγという3部品からなっているG蛋白からαサブユニットが解離する。
解離したαはフォスフォジエステラーゼ という酵素を活性化し、cGMPを分解する。
その結果イオンチャネルが閉じ、過分極を起こす。
http://www.ccn.yamanashi.ac.jp/~k-sakai/visual.html
***参考資料***
制御様式
イオンチャネルの開閉の制御様式には、いくつかある。
電位依存性 (voltage-dependent channel/voltage-gated channel)
:膜電位の変化に応じて開くもの。
時定数の異なる複数のゲートを持ち、
膜電位変化時に時間に依存した特定の開閉を行うチャネルも多い。
リガンド依存性(ligand-gated channel):特異的に結合する分子によるもの。
この場合イオンチャネル自体が受容体となっている。受容体の側から見れば、
イオンチャネル共役型受容体とも呼べる。
たとえば、AMPA型グルタミン酸受容体、NMDA型グルタミン酸受容体など。
リン酸化依存性:他タンパクからのリン酸化シグナルによるもの
力学的変形依存性:チャネル分子に機械的変形が関わると開くもの。
内耳の有毛細胞などが有名。
温度依存性:温度によるもの。
漏洩チャネル:通常開いており、少しずつ特定のイオンを漏らすように流すもの。
http://ja.wikipedia.org/wiki/%E3%82%A4%E3%82%AA%E3%83%B3%E3%83%81%E3%83%A3%E3%83%8D%E3%83%AB
活動電位からシナプス電位へ
シナプスにおいて、活動電位からシナプス電位に変換されます(図5)。
活動電位は、神経細胞の軸索突起の根元で発生し、軸索をつたわってシナプス前部に達します。
シナプス前部には膜電位感受性カルシウムチャネルがありますので、
これが活動電位によって膜電位が上昇したことを感知して、活性化し、
つまりチャネルが開いてカルシウムが細胞内にはいります。
このカルシウムによって、シナプス前部にあったシナプス小胞が細胞膜に融合し、
中に入っていた神経伝達物質をシナプス間隙に放出します。
興奮性の神経伝達を司っているグルタミン酸を例にあげてみますと、
グルタミン酸がシナプス後膜にあるその受容体、つまりグルタミン酸受容体に結合します。
グルタミン酸受容体は4つの膜蛋白からなり、それらのまん中に穴(チャネル)を形成していて、
受容体にグルタミン酸が結合するとタンパク質の構造が変化して、短時間穴が開きます。
この穴はカチオン(陽イオン;ナトリウム、カリウム、カルシウムなど)をとおす性質があるため、
細胞外には内側に比べて高濃度のナトリウムイオンがあり、
それがチャネルをとおって細胞内に流入します。ナトリウムイオンはプラスですから、
その結果、細胞内のマイナスの程度が浅くなる、つまり脱分極します。
脱分極するにつれ、カリウムイオンの流出もあるために、
活動電位のように膜電位がプラスまで上昇することはありません。
このような神経伝達物質によって引き起されたシナプス後部の膜電位をシナプス電位といいます。
グルタミン酸 (Glu) の場合、シナプス電位は脱分極性なので興奮性シナプス後電位といいます。
一方、GABA (ギャバ) やグリシンは抑制性です。
抑制性ニューロンからGABAが分泌されると、GABA受容体チャネルが活性化し、
塩素イオンを選択的に通す穴がひらき、細胞外に多い塩素イオンが細胞内に流入します。
塩素イオンはマイナスなので、その結果膜電位はよりマイナスの方向に深くなり、
つまり過分極になり、これを抑制性シナプス後電位といいます。
これによって、活動電位が出にくくなります(図5)。
麻酔薬、睡眠薬、抗不安薬などはこのGABA受容体の働きを促進させ、
グルタミン酸による興奮性の神経活動を抑制させるのです。
http://www.tmin.ac.jp/medical/17/synapse2.html
AMPA型グルタミン酸受容体
電流-電圧特性
GluR2を持つ受容体の他の相違点としては、比較的オームの法則に従った、
線形の電流-電圧特性を持つことがあげられる。
他のサブユニットのみで構成されるAMPA受容体は、
膜電位が負の状態ではオームの法則に従うが、正の状態ではほとんど電流を流さない、
内向きの整流性を持つことが知られている(右図参照)。
これは、膜電位が正のとき、GluR1,3,4の各サブユニットは
細胞内ポリアミンによる阻害を受けているためである。
http://ja.wikipedia.org/wiki/AMPA%E5%9E%8B%E3%82%B0%E3%83%AB%E3%82%BF%E3%83%9F%E3%83%B3%E9%85%B8%E5%8F%97%E5%AE%B9%E4%BD%93
NMDA型グルタミン酸受容体
NMDA受容体は通常不活性な性質を持つ。
これは、細胞外からのマグネシウムイオンがこの受容体の活動を阻害しているためである
(マグネシウム・ブロック)。ただし、これは膜電位が大きく負の時に限られている。
そのため、膜電位が正であるか、-10~-20 mV 程度の範囲においては
マグネシウムイオン阻害がかからず、
結果として受容体の電気生理学的特性はオームの法則に従う。
膜電位が大きく負になるとマグネシウムイオンの阻害がかかり始め、
-60~-70 mV 程度の静止膜電位に相当する膜電位では、ほとんど電流を流さない。
これらのことから、NMDA型受容体は外向き整流性を持つ、と称される。
また、刺激に応じて流す電流は、AMPA受容体に比べて遅く、持続的である。
http://ja.wikipedia.org/wiki/NMDA%E5%9E%8B%E3%82%B0%E3%83%AB%E3%82%BF%E3%83%9F%E3%83%B3%E9%85%B8%E5%8F%97%E5%AE%B9%E4%BD%93
5、シナプスのイオンチャネル
(1)活動電位を発生させるイオンチャネル=電位依存性チャネル
シナプス電位に応じてチャネルを開閉しており以下の2種類ある
①細胞内にイオンが流入してプラス電位を発生させるNa+チャネル
②細胞外にイオンが流出してマイナス電位を発生させるK+チャネル
(2)通常の神経細胞のシナプス電位を発生させるイオンチャネル
=リガンド依存性チャネルと漏洩チャネル
(2-1)リガンド依存性チャネル
脳内において神経細胞のシナプス間を神経伝達物質が伝導して、神経伝達物質によって
シナプス内に電位を発生させているイオンチャネルは、
リガンド依存性チャネルであり、以下の2種類がある
①プラス電位を発生させるNa+とK+チャネル
これは1つのチャネルでNa+とK+の2つを通すということである
②Cl-チャネル
Cl-チャネルは、静止膜電位と同じ電位を発生させる
つまり、プラス電位を発生させるのを抑制するような働きがある
(2-2)漏洩チャネル
シナプスには、K+とCl-のイオンが通れる漏洩チャネルがあり
常にK+イオンとCl-イオンが細胞内外を、
イオン濃度と膜電位に応じて自由に移動している
この漏洩チャネルが-70mvの静止膜電位を発生させている原因となっている
つまり、イオンポンプが
3個のナトリウムイオン(Na+)を細胞外に汲み出すと共に、
細胞内に2個のK+イオンを流入させている一方で、
K+の漏洩チャネルによりK+イオンが細胞外へと流出させている
以下にその関係を単純化させた式を示す
細胞内の電荷
=+2(イオンポンプによる2個のK+流入)
-3(イオンポンプによる3個のNa+流出)
-1(K+の漏洩チャネルによりK+流出)
=-2
つまりイオンポンプだけなら、-70mvよりも高い電位であったろうが
K+の漏洩チャネルによりK+イオンが流出して-70mvになっている
***参考資料***
漏洩チャネルの貢献
神経細胞の典型的な軸索において、静止膜電位は負であり、おおよそ-70mV程度である。
このことは、細胞外に陽イオンが比較的多い(もしくは細胞内に陰イオンが比較的多い)
ことを示唆している。実際には前者が正しい。
前述したNa+-K+交換イオンポンプは3個のナトリウムイオンと2個のカリウムイオンを
交換しているだけなので膜電位の変化にはそれほど大きく寄与しないが、
外にくみ出されたナトリウムイオンが細胞内に入り込むための
ナトリウムチャネルは通常不活性化されており、
開いていないのに対し、カリウムが細胞外に流出するカリウムチャネルの中には、
通常開きっぱなしのものが存在する。つまり、カリウムイオンは汲み入れても汲み入れても、
ある程度は細胞外に漏れ出て行ってしまうのである。
これが、静止膜電位が負になってしまう主な原因である。
このカリウムイオンを漏れ出させてしまうチャネルを、カリウム漏洩チャネルと呼ぶ。
http://ja.wikipedia.org/wiki/%E8%86%9C%E9%9B%BB%E4%BD%8D
塩化物イオン(Cl-)の移動は細胞膜を通してかなり自由度が高いため、
この膜電位による移動にほとんど依存している。
http://ja.wikipedia.org/wiki/%E8%86%9C%E9%9B%BB%E4%BD%8D
6、イオンチャネルを流れるイオン電流
イオンを流す方向は、チャネルによって決める事はできない。
なぜなら、イオンチャネルはイオンを受動的に輸送するからである。
イオンチャネルが開くと、イオンは以下の電流が流れる
(1)膜内外のイオンの濃度差により濃度を平均化しようとして流れる拡散電流
(2)膜電位差による電圧電流
***参考資料***
イオンの選択性はチャネルによってまちまちであり、
一種類のイオンのみ選択的に透過させるチャネルもあれば、
多くの陽イオンを通してしまうチャネルも存在する。
しかし、イオンを流す方向は、チャネルによって決める事はできない。
なぜなら、イオンチャネルはイオンを受動的に輸送するからである。
イオンチャネルが開くと、イオンは電気化学的勾配の高い方から低い方へ流れる。
つまり、イオンチャネルがいくら開いても、膜電位が平衡電位と等しい場合、
すなわち、膜内外でのイオン濃度差による拡散の効果と、
膜電位によるクーロンポテンシャルの効果が
釣り合っているとイオン電流は流れない。
http://ja.wikipedia.org/wiki/%E3%82%A4%E3%82%AA%E3%83%B3%E3%83%81%E3%83%A3%E3%83%8D%E3%83%AB
イオンチャネルによるイオンの移動
次に、イオンポンプなどの活動により一旦イオン分布の差が生まれると、
今度はその濃度差を利用した受動輸送が可能になる。
この受動輸送は、イオンチャネルと呼ばれるタンパク質によってなされる。
イオンチャネルは、イオンポンプ等によって濃度差が作られたイオンを、
イオン濃度の高いほうから低いほうへ拡散させる、イオンの通り道である。
よって、方向に選択性はなく、
膜電位がない場合は常にイオンの濃度勾配に従った輸送である。
ただし、イオン濃度の低い方から高い方への移動が
全くないわけではないことに注意すべきである。
イオンがチャネルを通過するかどうかはそのイオンがブラウン運動によって
チャネル分子に衝突するかどうかに依存しており、イオン濃度の高い側では、
イオンのチャネルへの衝突が、低いほうに比べて圧倒的に起きやすいため、
全体としては高い方から低い方への流れが生じるわけである。
イオンチャネルの多くは通常不活性型であり、
何らかの刺激(膜電位の変化・リガンドの結合・リン酸化・機械刺激など)に
応じて開閉する。
そのため、定常状態の細胞において、
働いているイオンチャネルは少ないと言える。
ただし、漏洩チャネルと呼ばれるタイプのイオンチャネルは常に開いており、
静止膜電位に貢献する(後述)。
膜電位そのものによるイオンの移動
膜内外の電位差そのものも、イオンの移動に影響を及ぼす。
たとえば、静止状態の膜電位は細胞内が細胞外に比べて
負であることはすでに述べたが、
負の細胞内に向けて陽イオンは入りやすく、陰イオンは入りにくい。
逆に、正の細胞外に向けて陰イオンは出て行きやすく、陽イオンは出て行きにくい。
これは、単純に細胞外の正電荷を持つ環境が、陽イオンを反発させようとするからである。
現実に、塩化物イオン(Cl-)の移動は細胞膜を通してかなり自由度が高いため、
この膜電位による移動にほとんど依存している。
http://ja.wikipedia.org/wiki/%E8%86%9C%E9%9B%BB%E4%BD%8D
7、平衡電位と拡散電位
(1)平衡電位と拡散電位の説明
膜を通して流れる電流 = 拡散電流+電圧電流 = 0
となっている状態を平衡状態という
つまり平衡状態とは、
ある量Aと別のある量Bが一致して平衡しているときの状態をいう
この平衡状態では、拡散電流を阻止するように電圧電流が逆に流れているので
見かけ上は正味の電流は流れていない
この拡散電流と平衡する電圧電流の膜電位を、平衡電位といい
各イオンごとによって平衡電位が存在する
この平衡電位は、拡散電流を電圧電流と考えたときの電圧に相当するので、
拡散電位ともいう
ただし拡散電位というときは、
拡散電流(拡散電位電流) = -電圧電流(平衡電位電流)
なので平衡電位の逆の符号の電位となる
電気回路理論で、拡散電流を取り扱うときは
この平衡電位(拡散電位)の電圧(一般には化学電池と呼ばれている)
によって電流が流れるとして取り扱う
つまり、各イオンごとの平衡電位(拡散電位)が
拡散電流を流す電圧として考えることができるということである
(2)平衡電位の求め方(ネルンストの式)
各イオンごとの平衡電位の求め方は、以下のネルンストの式で求められる
平衡電位 : Eion
ただし、lnは自然対数(eを底とする対数)であり、他のパラメータは以下のとおりである。
* R=気体定数(8.31J/mol/K)
* T=絶対温度(℃+273)
* z=イオンの荷数
* F=ファラデー定数(1molあたりの電荷、96500クーロン/mol)
* [ion]o=細胞外のイオン濃度
* [ion]i=細胞内のイオン濃度
それでカリウム、ナトリウム、塩素イオンの平衡電位を求めると以下のようになる
カリウムイオン(K+)の平衡電位 :Ek=-88.27[mV]
ナトリウムイオン(Na+)の平衡電位:Ena=58.67[mV]
塩素(Cl-)の平衡電位:Ecl=-70.25[mV]
(3)ネルンストの式の導出
ネルンストの式の導出は、平衡状態においては
個々のイオンの濃度が時間によって一定で変化がないので
それぞれのイオンに対して
個々のイオンが膜を通して流れる電流 = 拡散電流+電圧電流 = 0
とおいて計算することにより個々のイオンの平衡電位を求めている
しかし、実際は、
個々のイオンが膜を通して流れる電流
= 拡散電流+電圧電流+イオンポンプによる電流 = 0
が正しい計算式である
なので多少の誤差などが存在していると思われるが、
イオンポンプによる電流が拡散電流や電圧電流による電流よりも
無視できるほど小さいなら問題はない
(4)絶対温度が0[K](-273℃)で平衡電位が0Vになる理由
ネルンストの式から平衡電位は、
絶対温度が0[K](-273℃)になると0となる
このことを理解するためには拡散電流がどのようにして
流れるのかを理解する必要がある
拡散電流というのは熱エネルギーによってランダムな運動をする熱運動
(ブラウン運動)によって確率的に膜にぶつかり、
濃度が高ければ高いほど膜にあたるイオンの数が多くなるので、
両方とも同じ温度なら濃度の高い方から濃度の低い方へと拡散電流が流れるものである
そのため、絶対温度が0[K](-273℃)になると
熱運動をしなくなるので拡散電流は0となる
(実際は、量子力学的な揺らぎによって0点振動を起こしているので
完全に止まることはない
参照:固体物理学
http://163.51.52.40/users/kondo/lectures/ssp2007.pdf)
拡散電流が0となると、膜電位による電圧電流だけ流れることになり
体温での静止膜電位は-70mVなので、細胞外から細胞内へと
電圧電流が流れる
膜を通して流れる電流 = 膜電位による電圧電流
細胞内から細胞外へと流れる拡散電流がないので
細胞外から細胞内へと流れる膜電位による電圧電流によって
膜電位がどんどん減少していき、0Vになるまで電圧電流が流れることになる
したがって、絶対温度が0[K](-273℃)になると
平衡電位は0Vになるというこうことであり、
膜電位も当然0Vにならざるえないということである
(0Vよりも膜電位があればすぐに電圧電流が流れて0Vになる)
(これはイオンチャネルが絶対温度が0[K](-273℃)でも
開いていると仮定していることに注意する必要がある)
ここでもし、イオンポンプが絶対温度が0[K](-273℃)でも動作すると
仮定するならば(実際には生体細胞が0[K](-273℃)で動作することはない)
膜を通して流れる電流 = 電圧電流+イオンポンプによる電流 = 0
となる電圧で平衡することになる
(5)膜電位の求め方(ゴールドマンの式)
ネルンストの式では、個々のイオンの平衡電位を求めることはできても
複数のイオンによる平衡状態の膜電位を求めることはできない
平衡状態の膜電位(静止膜電位)は、以下のゴールドマンの式で求められる
ただし、lnは自然対数(eを底とする対数)であり、他のパラメータは以下のとおりである。
* R=気体定数(8.31J/mol/K)
* T=絶対温度(℃+273)
* z=イオンの荷数
* F=ファラデー定数(1molあたりの電荷、96500クーロン/mol)
* [ion]o=細胞外のイオン濃度
* [ion]i=細胞内のイオン濃度
それでカリウム、ナトリウム、塩素イオンの平衡電位を求めると以下のようになる
カリウムイオン(K+)の平衡電位 :Ek=-88.27[mV]
ナトリウムイオン(Na+)の平衡電位:Ena=58.67[mV]
塩素(Cl-)の平衡電位:Ecl=-70.25[mV]
(3)ネルンストの式の導出
ネルンストの式の導出は、平衡状態においては
個々のイオンの濃度が時間によって一定で変化がないので
それぞれのイオンに対して
個々のイオンが膜を通して流れる電流 = 拡散電流+電圧電流 = 0
とおいて計算することにより個々のイオンの平衡電位を求めている
しかし、実際は、
個々のイオンが膜を通して流れる電流
= 拡散電流+電圧電流+イオンポンプによる電流 = 0
が正しい計算式である
なので多少の誤差などが存在していると思われるが、
イオンポンプによる電流が拡散電流や電圧電流による電流よりも
無視できるほど小さいなら問題はない
(4)絶対温度が0[K](-273℃)で平衡電位が0Vになる理由
ネルンストの式から平衡電位は、
絶対温度が0[K](-273℃)になると0となる
このことを理解するためには拡散電流がどのようにして
流れるのかを理解する必要がある
拡散電流というのは熱エネルギーによってランダムな運動をする熱運動
(ブラウン運動)によって確率的に膜にぶつかり、
濃度が高ければ高いほど膜にあたるイオンの数が多くなるので、
両方とも同じ温度なら濃度の高い方から濃度の低い方へと拡散電流が流れるものである
そのため、絶対温度が0[K](-273℃)になると
熱運動をしなくなるので拡散電流は0となる
(実際は、量子力学的な揺らぎによって0点振動を起こしているので
完全に止まることはない
参照:固体物理学
http://163.51.52.40/users/kondo/lectures/ssp2007.pdf)
拡散電流が0となると、膜電位による電圧電流だけ流れることになり
体温での静止膜電位は-70mVなので、細胞外から細胞内へと
電圧電流が流れる
膜を通して流れる電流 = 膜電位による電圧電流
細胞内から細胞外へと流れる拡散電流がないので
細胞外から細胞内へと流れる膜電位による電圧電流によって
膜電位がどんどん減少していき、0Vになるまで電圧電流が流れることになる
したがって、絶対温度が0[K](-273℃)になると
平衡電位は0Vになるというこうことであり、
膜電位も当然0Vにならざるえないということである
(0Vよりも膜電位があればすぐに電圧電流が流れて0Vになる)
(これはイオンチャネルが絶対温度が0[K](-273℃)でも
開いていると仮定していることに注意する必要がある)
ここでもし、イオンポンプが絶対温度が0[K](-273℃)でも動作すると
仮定するならば(実際には生体細胞が0[K](-273℃)で動作することはない)
膜を通して流れる電流 = 電圧電流+イオンポンプによる電流 = 0
となる電圧で平衡することになる
(5)膜電位の求め方(ゴールドマンの式)
ネルンストの式では、個々のイオンの平衡電位を求めることはできても
複数のイオンによる平衡状態の膜電位を求めることはできない
平衡状態の膜電位(静止膜電位)は、以下のゴールドマンの式で求められる
[K+]o =カリウムイオンの細胞外のイオン濃度
[K+]i =カリウムイオンの細胞内のイオン濃度
[Na+]o =ナトリウムイオンの細胞外のイオン濃度
[Na+]i =ナトリウムイオンの細胞内のイオン濃度
[Cl-]o =塩化物イオンの細胞外のイオン濃度
[Cl-]i =塩化物イオンの細胞内のイオン濃度
Pk、Pna、Pclは、それぞれのイオンの透過係数で
生体内においてはおおよそPK : PNa : PCl = 1 : 0.04 : 0.45程度である
ゴールドマンの式で、平衡状態の静止膜電位を計算すると
-70mVとなり実験値とほとんど一致する
静止膜電位: Em=-70[mV]
(6)ゴールドマンの式の導出
ゴールドマンの式の導出は、平衡状態においては膜を通して流れる電流が0であり、
膜を通ることができるイオンをNa+、K+、Cl-の3つだけと考えて以下のように式をたてた
膜を通して流れる電流 =
Na+の拡散電流と電圧電流 +
K+の拡散電流と電圧電流 +
Cl-の拡散電流と電圧電流
= 0
さらに仮定として電位勾配:φが
dφ/dx = E/a
x:膜間距離
E:膜の電界
a:膜の厚み
のように一定であるとして、変数を一つ減らすことにより
解析的に求めた結果である
式の導出方法からわかるように、イオンポンプの電流を無視しているのだが
導かれた結果は正確に実験データと一致するので
イオンポンプによる電流はそれほど問題にならないほど小さいと推測される
(7)膜電位の求め方(等価回路方式)
ゴールドマンの式を導出するのは少し大変であり
もっと簡単に膜電位を求める方法がある
膜の平衡状態を、Na+、K+、Cl-の平衡電位(拡散電位)とその抵抗成分などを用いて
等価回路で表現し、その等価回路の出力電圧を求める方法で計算する方法である
この等価回路方式で静止膜電位を計算すると、だいたい-78.8mVとなり、
少し値の一致がよくないがそれほど悪くはない結果となる
(等価回路方式による膜電位の求め方は、別の記事で詳細に解説します)
ここで重要なことは、ゴールドマンの式だと
平衡状態の電流が流れてないときの膜電位なら求められるが
非平衡状態の電流が流れているときの膜電位を計算することができないことである
つまり、ゴールドマンの式からだと
膜電位がどのように変化するのかわからないというという欠点がある
それに対して、等価回路方式を用いて膜電位を求めると、多少の誤差はあるが
(誤差の原因はNa+、K+、Cl-の抵抗成分が正確に求められてないことにあると推測される
それを正確に求めれば完璧に一致する可能性が高い)、
かなり確かな値が求まり、しかも時間によって膜電位がどのように変化するか
計算することができるという素晴らしい点がある
実際に等価回路によって後に述べる活動電位の波形を
極めて正確に説明することができるので、
等価回路によって正確に膜電位の変化、値を求めることができるということである
このように等価回路によって表現することは非常に重要であり、
別の記事で神経細胞を等価回路によって正確に表現し、
その回路から回路方程式をたてて
神経細胞内の電気信号を正確に計算する方法を示します
***参考資料***
平衡電位、ネルンストの式、ゴールドマンの式
http://ja.wikipedia.org/wiki/%E8%86%9C%E9%9B%BB%E4%BD%8D
ネルンストの式とゴールドマンの式の導出方法
http://user.ecc.u-tokyo.ac.jp/~ckam/Nernst_equation.pdf
<<活動電位、シナプス電位、化学信号について>>
1、シナプス間の結合の仕方:化学シナプスと電気シナプス
(1)化学シナプス=シナプス間を神経伝達物質の化学信号で信号を伝達するシナプス結合
[K+]i =カリウムイオンの細胞内のイオン濃度
[Na+]o =ナトリウムイオンの細胞外のイオン濃度
[Na+]i =ナトリウムイオンの細胞内のイオン濃度
[Cl-]o =塩化物イオンの細胞外のイオン濃度
[Cl-]i =塩化物イオンの細胞内のイオン濃度
Pk、Pna、Pclは、それぞれのイオンの透過係数で
生体内においてはおおよそPK : PNa : PCl = 1 : 0.04 : 0.45程度である
ゴールドマンの式で、平衡状態の静止膜電位を計算すると
-70mVとなり実験値とほとんど一致する
静止膜電位: Em=-70[mV]
(6)ゴールドマンの式の導出
ゴールドマンの式の導出は、平衡状態においては膜を通して流れる電流が0であり、
膜を通ることができるイオンをNa+、K+、Cl-の3つだけと考えて以下のように式をたてた
膜を通して流れる電流 =
Na+の拡散電流と電圧電流 +
K+の拡散電流と電圧電流 +
Cl-の拡散電流と電圧電流
= 0
さらに仮定として電位勾配:φが
dφ/dx = E/a
x:膜間距離
E:膜の電界
a:膜の厚み
のように一定であるとして、変数を一つ減らすことにより
解析的に求めた結果である
式の導出方法からわかるように、イオンポンプの電流を無視しているのだが
導かれた結果は正確に実験データと一致するので
イオンポンプによる電流はそれほど問題にならないほど小さいと推測される
(7)膜電位の求め方(等価回路方式)
ゴールドマンの式を導出するのは少し大変であり
もっと簡単に膜電位を求める方法がある
膜の平衡状態を、Na+、K+、Cl-の平衡電位(拡散電位)とその抵抗成分などを用いて
等価回路で表現し、その等価回路の出力電圧を求める方法で計算する方法である
この等価回路方式で静止膜電位を計算すると、だいたい-78.8mVとなり、
少し値の一致がよくないがそれほど悪くはない結果となる
(等価回路方式による膜電位の求め方は、別の記事で詳細に解説します)
ここで重要なことは、ゴールドマンの式だと
平衡状態の電流が流れてないときの膜電位なら求められるが
非平衡状態の電流が流れているときの膜電位を計算することができないことである
つまり、ゴールドマンの式からだと
膜電位がどのように変化するのかわからないというという欠点がある
それに対して、等価回路方式を用いて膜電位を求めると、多少の誤差はあるが
(誤差の原因はNa+、K+、Cl-の抵抗成分が正確に求められてないことにあると推測される
それを正確に求めれば完璧に一致する可能性が高い)、
かなり確かな値が求まり、しかも時間によって膜電位がどのように変化するか
計算することができるという素晴らしい点がある
実際に等価回路によって後に述べる活動電位の波形を
極めて正確に説明することができるので、
等価回路によって正確に膜電位の変化、値を求めることができるということである
このように等価回路によって表現することは非常に重要であり、
別の記事で神経細胞を等価回路によって正確に表現し、
その回路から回路方程式をたてて
神経細胞内の電気信号を正確に計算する方法を示します
***参考資料***
平衡電位、ネルンストの式、ゴールドマンの式
http://ja.wikipedia.org/wiki/%E8%86%9C%E9%9B%BB%E4%BD%8D
ネルンストの式とゴールドマンの式の導出方法
http://user.ecc.u-tokyo.ac.jp/~ckam/Nernst_equation.pdf
<<活動電位、シナプス電位、化学信号について>>
1、シナプス間の結合の仕方:化学シナプスと電気シナプス
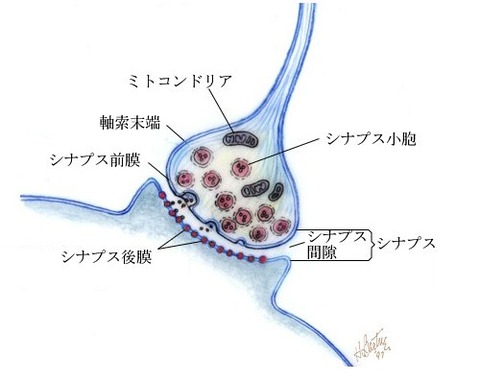
(1)化学シナプス=シナプス間を神経伝達物質の化学信号で信号を伝達するシナプス結合


化学シナプスとは、細胞間に神経伝達物質が放出され、
それがシナプスの受容体に結合することによって細胞間の
情報伝達が行われるシナプスのことを指す。
化学シナプスにおける神経細胞(ニューロン)のシナプスとシナプスの間には、
20~30ナノメートル程度の間隙があり
その間隙を神経伝達物質(脳内ホルモンともいう)の化学物質がやり取りされて
ニューロン間で信号が伝達する
この神経伝達物質の化学物質による信号が、化学信号である
化学シナプスは、神経細胞の前シナプス(活動電位が伝わる終端)と
別の神経細胞の後シナプス(活動電位を発生させる部分)を結合させている
化学シナプスでは、シナプス間で信号を伝達するのに
0.3から1ミリ秒(ミリ秒は1000分の1秒)の遅れ時間がある
参照:化学シナプスにおける神経伝達画像
(2)電気シナプス=シナプス間をイオン電流の電気信号で信号を伝達するシナプス結合
電気シナプスとは、細胞間がイオンなどを通過させる分子で接着され、
細胞間に直接イオン電流が流れることによって
細胞間のシグナル伝達が行われるシナプスのことを指す
電気シナプスは、シナプス間が2ナノメートル程度のギャップしかなく、
シナプス間でイオン電流の電気信号を伝達するこにより信号を伝達する
化学信号に比較して非常に高速に信号を伝達させることができる
電気シナプスは化学シナプスと違い、
神経細胞の後シナプス(活動電位を発生させる部分)と
別の神経細胞の後シナプス(活動電位を発生させる部分)とを主につなげている
参照:化学シナプスと電気シナプスにおける神経伝達画像
***参考資料***
(1)化学シナプス
化学シナプスとは、細胞間に神経伝達物質が放出され、
それが受容体に結合することによって細胞間の情報伝達が
行われるシナプスのことを指す。
化学シナプスは電気シナプスより広範に見られ、
一般にシナプスとだけ言われるときはこちらを指すことが多い。
構造と機序
化学シナプスの基本的構造は、
神経細胞の軸索の先端が他の細胞(神経細胞の樹状突起や筋線維)と
20nm程度の隙間(シナプス間隙)を空けて、シナプス接着分子によって
細胞接着している状態である。
情報伝達は一方向に行われ、興奮がシナプスに達するとシナプス小胞が
細胞膜に融合しシナプス間隙に神経伝達物質が放出される。
そして拡散した神経伝達物質がシナプス後細胞に存在する
受容体に結合することで刺激が伝達されて行く。
化学シナプスにおける典型的な情報伝達機序は以下のように進む。
1. 前シナプス細胞の軸索を活動電位が伝わり、末端にある膨らみであるシナプス小頭に到達する
2. 活動電位によりシナプス小頭の膜上に位置する電位依存性カルシウムイオンチャネルが開く。
3. するとカルシウムイオンがシナプス内に流入し、シナプス小胞が細胞膜に接して神経伝達物質が細胞外に開口放出される。
4. 神経伝達物質はシナプス間隙を拡散し、後シナプス細胞の細胞膜上に分布する神経伝達物質受容体に結合する。
5. 後シナプス細胞のイオンチャネルが開き、細胞膜内外の電位差が変化する。
(2)電気シナプス
電気シナプスとは、細胞間がイオンなどを通過させる分子で接着され、
細胞間に直接イオン電流が流れることによって細胞間のシグナル伝達が行われるシナプスのことを指す。
網膜の神経細胞間や心筋の筋繊維間などで広範に見られる。
化学シナプスのように方向づけられた伝達はできないが、それよりも高速な伝達が行われ、
多くの細胞が協調して動作する現象を引き起こす。
電気シナプスは無脊椎動物の神経系では一般的にみられるが、長らく脊椎動物の中枢神経系では見出されておらず、
脊椎動物の脳での神経伝達は化学シナプスのみによるものと考えられていた。
後になって海馬や大脳皮質の抑制性介在神経細胞の樹状突起間で発見され、
重要な伝達手段となっていることが見出された。
http://ja.wikipedia.org/wiki/%E3%82%B7%E3%83%8A%E3%83%97%E3%82%B9
化学シナプスと電気シナプス
シナプス部分に、ギャップがあるかどうかという論争は、ギャップがあるということで、
ひとまず決着したが、その後、原形質どおしが連絡した、
電気シナプスと呼ばれるタイプのものが見つかった。
化学シナプスでは、細胞膜どうしは完全に離れており、その間に、
20ないし30ナノメートル(1ナノメートルは、10万分の1ミリメートル)のギャップがあった。
電気シナプスでは、細胞膜の間に2ナノメートル程度のギャップしかなく、
しかも、2つの神経細胞の細胞膜が細い管を作って融合し、その管の通じて原形質が連絡している。
この管が、シナプス部分全体に規則的に分布している。原形質が融合しているために、
連絡しあう2つの神経細胞のどちらかに電位変化が起こると、イオンの流れとして、
他の神経細胞へと電位変化が伝わる。そのため、シナプス部分での信号伝達の遅れ時間は非常に小さい。
化学シナプスでは、0.3から1ミリ秒(ミリ秒は1000分の1秒)の遅れ時間があるが、
電気シナプスでは遅れ時間は、ほとんどゼロである。
その一方、単に原形質が連絡しているだけなので、信号の伝達の方向は、双方向性である。
また、薬物の影響を受けにくく、信号の合成という点でも、殆ど役割を果たさない。
http://www.pri.kyoto-u.ac.jp/brain/brain/15-1/index-15-1.html
2、化学シナプスによるシナプス電位の変化
(1)Na+とK+を通すリガンド依存性チャネルにより発生するシナプス電位
シナプス電位を発生させるチャネルは、リガンド依存性チャネルで
Na+とK+を通すチャネルがある
これは1つのチャネルでNa+とK+を通すということである
神経伝達物質のような化学物質をチャネルの受容体で受け取るとチャネルが開く
このチャネルを通って流れるNa+とK+イオン電流は、以下のようになる
①拡散電流
拡散電流 = Na+拡散電流(拡散電位:-60mVによる細胞外から細胞内への電流)
+ K+拡散電流(拡散電位:90mVによる細胞内から細胞外への電流)
= 15mVの拡散電流
15mVによる拡散電流となるのは、
Na+とK+に対するイオンチャネルの抵抗の関係でこうなる
(この拡散電流の計算は、別の記事で計算します)
つまり、拡散電流はK+拡散電流成分の方が大きく、
細胞外へと流出する方に流れることになる
②電圧電流
電圧電流 = Na+とK+の電圧電流(膜電位の電圧電流)
③膜を流れる電流
膜を流れる電流=拡散電流+電圧電流=15mV+膜電位による電流
となり、イオンチャネルがいくら開いても
電圧電流が拡散電流(15mV)と平衡する
-15mVの膜電位までしかならないことがわかる
(2)シナプス電位の変化
他のニューロンから出た神経伝達物質の化学物質を受け取る後シナプス細胞内の電位を、
シナプス電位(またはシナプス後電位、後シナプス電位)という
シナプス電位は、シナプス内の電荷を持ったイオン
(膜電位の計算のときの説明から、Na+、K+、Cl-のイオン)により生じており
そのイオンの電荷量が変化することによってシナプス電位が変化する
シナプスは、化学物質を受け取る受容体があり、その受容体で化学物質を受け取ると
Na+とK+イオンを通過させるイオンチャネルが開き、(1)で述べたように
Na+とK+イオンの拡散電流と膜電位による電圧電流が流れて
静止膜電位より高いシナプス電位となる
イオンチャネルの項で説明したように、化学物質によりイオンチャネルが開閉するので
このようなチャネルをリガンド依存性チャネルという
化学物質の信号により一定時間、イオンチャネルが開いて
Na+とK+イオンのイオン電流が流れた後、
化学物質は受容体から放出され、イオンチャネルは閉じる
放出後は、化学物質は速やかに酵素によって不活性化されるか、
または前シナプス終末に再吸収され、一部は再び再利用される
シナプスは、イオンポンプによる細胞外へのNa+イオンの放出と、
漏洩チャネルを通して細胞外へのK+イオンの流出
(K+の拡散電位より膜電位が低くなっているのでK+イオンが拡散により細胞外へと流出する)
によって常に余分に流入したイオンを排出しており静止電位へと戻ろうとする
なので、化学物質を受け取り一定時間Na+とK+イオン電流が流れることによって
発生したシナプス電位は
イオンポンプと漏洩チャネルによる細胞外へのNa+とK+イオンの流出により
時間が経つと静止膜電位へと戻る
静止状態でのシナプス電位は、静止膜電位なので、
およそ-70mVである(細胞外の電位を0としている)。
以上のように神経細胞(ニューロン)のシナプスの受容体部分に、
神経伝達物質の化学信号が伝達すると
それに応じてシナプス内に、電荷を持ったイオンの流入が起こり、
シナプス内部の電位(シナプス電位)が変化します(=電気信号発生)
1つの化学信号は一定の時間だけイオン電流を細胞内へと流し
また常にイオンポンプと漏洩チャネルにより
細胞内のプラス電荷(Na+とK+)を細胞外へと流出させて静止電位へと戻ろうとするので
この化学信号によって生じたイオン電荷の変動による電位変化は一時的なものであり、
時間が経てばすぐに元の電位(約-70mV)に戻る
また、シナプスは、1個の神経細胞に100個から10万個あり、
(おそらく前シナプス、後シナプスの入出力あわせての数なので、
どちらか一方のシナプスの入力だけなら50個から5万個ぐらい)
数千から数万もの化学信号の入力をうけている
それがシナプスの受容体に結合することによって細胞間の
情報伝達が行われるシナプスのことを指す。
化学シナプスにおける神経細胞(ニューロン)のシナプスとシナプスの間には、
20~30ナノメートル程度の間隙があり
その間隙を神経伝達物質(脳内ホルモンともいう)の化学物質がやり取りされて
ニューロン間で信号が伝達する
この神経伝達物質の化学物質による信号が、化学信号である
化学シナプスは、神経細胞の前シナプス(活動電位が伝わる終端)と
別の神経細胞の後シナプス(活動電位を発生させる部分)を結合させている
化学シナプスでは、シナプス間で信号を伝達するのに
0.3から1ミリ秒(ミリ秒は1000分の1秒)の遅れ時間がある
参照:化学シナプスにおける神経伝達画像
(2)電気シナプス=シナプス間をイオン電流の電気信号で信号を伝達するシナプス結合
電気シナプスとは、細胞間がイオンなどを通過させる分子で接着され、
細胞間に直接イオン電流が流れることによって
細胞間のシグナル伝達が行われるシナプスのことを指す
電気シナプスは、シナプス間が2ナノメートル程度のギャップしかなく、
シナプス間でイオン電流の電気信号を伝達するこにより信号を伝達する
化学信号に比較して非常に高速に信号を伝達させることができる
電気シナプスは化学シナプスと違い、
神経細胞の後シナプス(活動電位を発生させる部分)と
別の神経細胞の後シナプス(活動電位を発生させる部分)とを主につなげている
参照:化学シナプスと電気シナプスにおける神経伝達画像
***参考資料***
(1)化学シナプス
化学シナプスとは、細胞間に神経伝達物質が放出され、
それが受容体に結合することによって細胞間の情報伝達が
行われるシナプスのことを指す。
化学シナプスは電気シナプスより広範に見られ、
一般にシナプスとだけ言われるときはこちらを指すことが多い。
構造と機序
化学シナプスの基本的構造は、
神経細胞の軸索の先端が他の細胞(神経細胞の樹状突起や筋線維)と
20nm程度の隙間(シナプス間隙)を空けて、シナプス接着分子によって
細胞接着している状態である。
情報伝達は一方向に行われ、興奮がシナプスに達するとシナプス小胞が
細胞膜に融合しシナプス間隙に神経伝達物質が放出される。
そして拡散した神経伝達物質がシナプス後細胞に存在する
受容体に結合することで刺激が伝達されて行く。
化学シナプスにおける典型的な情報伝達機序は以下のように進む。
1. 前シナプス細胞の軸索を活動電位が伝わり、末端にある膨らみであるシナプス小頭に到達する
2. 活動電位によりシナプス小頭の膜上に位置する電位依存性カルシウムイオンチャネルが開く。
3. するとカルシウムイオンがシナプス内に流入し、シナプス小胞が細胞膜に接して神経伝達物質が細胞外に開口放出される。
4. 神経伝達物質はシナプス間隙を拡散し、後シナプス細胞の細胞膜上に分布する神経伝達物質受容体に結合する。
5. 後シナプス細胞のイオンチャネルが開き、細胞膜内外の電位差が変化する。
(2)電気シナプス
電気シナプスとは、細胞間がイオンなどを通過させる分子で接着され、
細胞間に直接イオン電流が流れることによって細胞間のシグナル伝達が行われるシナプスのことを指す。
網膜の神経細胞間や心筋の筋繊維間などで広範に見られる。
化学シナプスのように方向づけられた伝達はできないが、それよりも高速な伝達が行われ、
多くの細胞が協調して動作する現象を引き起こす。
電気シナプスは無脊椎動物の神経系では一般的にみられるが、長らく脊椎動物の中枢神経系では見出されておらず、
脊椎動物の脳での神経伝達は化学シナプスのみによるものと考えられていた。
後になって海馬や大脳皮質の抑制性介在神経細胞の樹状突起間で発見され、
重要な伝達手段となっていることが見出された。
http://ja.wikipedia.org/wiki/%E3%82%B7%E3%83%8A%E3%83%97%E3%82%B9
化学シナプスと電気シナプス
シナプス部分に、ギャップがあるかどうかという論争は、ギャップがあるということで、
ひとまず決着したが、その後、原形質どおしが連絡した、
電気シナプスと呼ばれるタイプのものが見つかった。
化学シナプスでは、細胞膜どうしは完全に離れており、その間に、
20ないし30ナノメートル(1ナノメートルは、10万分の1ミリメートル)のギャップがあった。
電気シナプスでは、細胞膜の間に2ナノメートル程度のギャップしかなく、
しかも、2つの神経細胞の細胞膜が細い管を作って融合し、その管の通じて原形質が連絡している。
この管が、シナプス部分全体に規則的に分布している。原形質が融合しているために、
連絡しあう2つの神経細胞のどちらかに電位変化が起こると、イオンの流れとして、
他の神経細胞へと電位変化が伝わる。そのため、シナプス部分での信号伝達の遅れ時間は非常に小さい。
化学シナプスでは、0.3から1ミリ秒(ミリ秒は1000分の1秒)の遅れ時間があるが、
電気シナプスでは遅れ時間は、ほとんどゼロである。
その一方、単に原形質が連絡しているだけなので、信号の伝達の方向は、双方向性である。
また、薬物の影響を受けにくく、信号の合成という点でも、殆ど役割を果たさない。
http://www.pri.kyoto-u.ac.jp/brain/brain/15-1/index-15-1.html
2、化学シナプスによるシナプス電位の変化
(1)Na+とK+を通すリガンド依存性チャネルにより発生するシナプス電位
シナプス電位を発生させるチャネルは、リガンド依存性チャネルで
Na+とK+を通すチャネルがある
これは1つのチャネルでNa+とK+を通すということである
神経伝達物質のような化学物質をチャネルの受容体で受け取るとチャネルが開く
このチャネルを通って流れるNa+とK+イオン電流は、以下のようになる
①拡散電流
拡散電流 = Na+拡散電流(拡散電位:-60mVによる細胞外から細胞内への電流)
+ K+拡散電流(拡散電位:90mVによる細胞内から細胞外への電流)
= 15mVの拡散電流
15mVによる拡散電流となるのは、
Na+とK+に対するイオンチャネルの抵抗の関係でこうなる
(この拡散電流の計算は、別の記事で計算します)
つまり、拡散電流はK+拡散電流成分の方が大きく、
細胞外へと流出する方に流れることになる
②電圧電流
電圧電流 = Na+とK+の電圧電流(膜電位の電圧電流)
③膜を流れる電流
膜を流れる電流=拡散電流+電圧電流=15mV+膜電位による電流
となり、イオンチャネルがいくら開いても
電圧電流が拡散電流(15mV)と平衡する
-15mVの膜電位までしかならないことがわかる
(2)シナプス電位の変化
他のニューロンから出た神経伝達物質の化学物質を受け取る後シナプス細胞内の電位を、
シナプス電位(またはシナプス後電位、後シナプス電位)という
シナプス電位は、シナプス内の電荷を持ったイオン
(膜電位の計算のときの説明から、Na+、K+、Cl-のイオン)により生じており
そのイオンの電荷量が変化することによってシナプス電位が変化する
シナプスは、化学物質を受け取る受容体があり、その受容体で化学物質を受け取ると
Na+とK+イオンを通過させるイオンチャネルが開き、(1)で述べたように
Na+とK+イオンの拡散電流と膜電位による電圧電流が流れて
静止膜電位より高いシナプス電位となる
イオンチャネルの項で説明したように、化学物質によりイオンチャネルが開閉するので
このようなチャネルをリガンド依存性チャネルという
化学物質の信号により一定時間、イオンチャネルが開いて
Na+とK+イオンのイオン電流が流れた後、
化学物質は受容体から放出され、イオンチャネルは閉じる
放出後は、化学物質は速やかに酵素によって不活性化されるか、
または前シナプス終末に再吸収され、一部は再び再利用される
シナプスは、イオンポンプによる細胞外へのNa+イオンの放出と、
漏洩チャネルを通して細胞外へのK+イオンの流出
(K+の拡散電位より膜電位が低くなっているのでK+イオンが拡散により細胞外へと流出する)
によって常に余分に流入したイオンを排出しており静止電位へと戻ろうとする
なので、化学物質を受け取り一定時間Na+とK+イオン電流が流れることによって
発生したシナプス電位は
イオンポンプと漏洩チャネルによる細胞外へのNa+とK+イオンの流出により
時間が経つと静止膜電位へと戻る
静止状態でのシナプス電位は、静止膜電位なので、
およそ-70mVである(細胞外の電位を0としている)。
以上のように神経細胞(ニューロン)のシナプスの受容体部分に、
神経伝達物質の化学信号が伝達すると
それに応じてシナプス内に、電荷を持ったイオンの流入が起こり、
シナプス内部の電位(シナプス電位)が変化します(=電気信号発生)
1つの化学信号は一定の時間だけイオン電流を細胞内へと流し
また常にイオンポンプと漏洩チャネルにより
細胞内のプラス電荷(Na+とK+)を細胞外へと流出させて静止電位へと戻ろうとするので
この化学信号によって生じたイオン電荷の変動による電位変化は一時的なものであり、
時間が経てばすぐに元の電位(約-70mV)に戻る
また、シナプスは、1個の神経細胞に100個から10万個あり、
(おそらく前シナプス、後シナプスの入出力あわせての数なので、
どちらか一方のシナプスの入力だけなら50個から5万個ぐらい)
数千から数万もの化学信号の入力をうけている

(3)一つの化学信号によってシナプスに発生する電気信号
一つの化学信号によってシナプスに発生する電位変化の電気信号は、
下記の図などから推測で以下のものである
http://www.biol.s.u-tokyo.ac.jp/users/naibunpi/Oka/2005Summer_Seminar.pdf
(3-1)電圧幅:約数mV~数十mV
(3-2)時間幅:約数十ms
(3-3)周波数:Min=数十Hz、MAX=数kHz程度
①周波数
周波数:fは、f=1/Tで求まり
信号波形の時間幅が約数十msなので、
周波数:f=1/T=1/(約数十ms)=数十Hz
となる
②MAX周波数:数kHz程度
信号波形の時間幅約数十ms~数百msとはいっても
その信号を周波数分析すると(フーリエ解析すると)、
ある一定の周波数帯域を持った信号であるのが普通
したがって、信号に高周波成分があるのは間違いない
また、信号の立ち上がりは急速に立ち上がるので
高周波成分となる
それがだいたい最大で数kHz程度(時間にして0.1~1ms)
ではないかと推測される
つまりシナプス電位の電気信号の最大周波数成分は、
だいたい数kHz程度ではないかと推測される
(3-4)波形
①立ち上がり
膜を流れる電流は先に述べたように以下の式で与えられる
膜を流れる電流=拡散電流+電圧電流=15mV+膜電位の電流
膜電位を静止膜電位とすると、静止膜電位は-70mVだから
膜を通過するNa+とK+によるイオン電流は
15mV-70mV =-55mV
の電圧電流となる
したがって、立ち上がりは高い電圧電流なので早く起こり、
電位波形の立ち上がりは早めに立ち上がる
②ピーク
イオンチャネルが閉じてNa+イオンの流入が終わった瞬間でピークになる
③立ち下り
イオンポンプと漏洩チャネルによりNa+、K+イオンが細胞外へと流出し
Cl-イオンが細胞内へと流入するので、電位波形はさがっていく
イオンポンプと漏洩チャネルによるNa+、K+、Cl-イオンの量の変化は、
静止膜電位の-70mVにしようとするので
70mVの拡散電位による細胞外への電流とみなせる
電圧電流は、膜電位がピーク時のシナプス電位による電圧電流である
なので膜を流れる電流は以下のようになる
膜を流れる電流
=70mV(拡散電位)による拡散電流+ピーク時のシナプス電位による電圧電流
(ピーク時のシナプス電位が-50mVだとすると、20mVの電圧)
したがって、立ち上がりのときに流れる電流:-55mVよりも
低い電流となるのでなだらかに下がっていくことになる
数式で表すと以下のような波形である
(数式の導出、正確な波形の求め方は別の記事で説明します)
一つの化学信号によってシナプスに発生する電気信号
=k{exp(-t/T1)-exp(-t/T2)}
T1>>T2
この波形の図を以下に示す
(3-1)電圧幅:約数mV~数十mV
(3-2)時間幅:約数十ms
(3-3)周波数:Min=数十Hz、MAX=数kHz程度
①周波数
周波数:fは、f=1/Tで求まり
信号波形の時間幅が約数十msなので、
周波数:f=1/T=1/(約数十ms)=数十Hz
となる
②MAX周波数:数kHz程度
信号波形の時間幅約数十ms~数百msとはいっても
その信号を周波数分析すると(フーリエ解析すると)、
ある一定の周波数帯域を持った信号であるのが普通
したがって、信号に高周波成分があるのは間違いない
また、信号の立ち上がりは急速に立ち上がるので
高周波成分となる
それがだいたい最大で数kHz程度(時間にして0.1~1ms)
ではないかと推測される
つまりシナプス電位の電気信号の最大周波数成分は、
だいたい数kHz程度ではないかと推測される
(3-4)波形
①立ち上がり
膜を流れる電流は先に述べたように以下の式で与えられる
膜を流れる電流=拡散電流+電圧電流=15mV+膜電位の電流
膜電位を静止膜電位とすると、静止膜電位は-70mVだから
膜を通過するNa+とK+によるイオン電流は
15mV-70mV =-55mV
の電圧電流となる
したがって、立ち上がりは高い電圧電流なので早く起こり、
電位波形の立ち上がりは早めに立ち上がる
②ピーク
イオンチャネルが閉じてNa+イオンの流入が終わった瞬間でピークになる
③立ち下り
イオンポンプと漏洩チャネルによりNa+、K+イオンが細胞外へと流出し
Cl-イオンが細胞内へと流入するので、電位波形はさがっていく
イオンポンプと漏洩チャネルによるNa+、K+、Cl-イオンの量の変化は、
静止膜電位の-70mVにしようとするので
70mVの拡散電位による細胞外への電流とみなせる
電圧電流は、膜電位がピーク時のシナプス電位による電圧電流である
なので膜を流れる電流は以下のようになる
膜を流れる電流
=70mV(拡散電位)による拡散電流+ピーク時のシナプス電位による電圧電流
(ピーク時のシナプス電位が-50mVだとすると、20mVの電圧)
したがって、立ち上がりのときに流れる電流:-55mVよりも
低い電流となるのでなだらかに下がっていくことになる
数式で表すと以下のような波形である
(数式の導出、正確な波形の求め方は別の記事で説明します)
一つの化学信号によってシナプスに発生する電気信号
=k{exp(-t/T1)-exp(-t/T2)}
T1>>T2
この波形の図を以下に示す
なお、各シナプスでの結合強度によって発生する
シナプス電位の電気信号は違う
以上のようなアナログ電気信号が
一つの化学信号によってシナプスに発生する電気信号であり、
複数の化学信号が入力されると、上記の電気信号が重ね合わさったような
アナログ電気信号となる
***参考資料***
参考文献:「生体電気信号とはなにか」 杉晴夫
活動電位とシナプス電位
脳の活動は活動電位とシナプス電位によって成り立っています。
活動電位は細胞体からシナプス前部まで軸索突起(アクソン)を伝わります。
活動電位が軸索の終末にくるとシナプス前部から神経伝達物質が分泌され、
それが樹状突起(デンドライト)のシナプス後部の受容体を活性化し、
シナプス後細胞にシナプス電位を発生させます。
神経細胞は数千から数万ものシナプス入力をうけており、
http://www.tmin.ac.jp/medical/17/synapse2.html
情報伝達は一方向に行われ、興奮がシナプスに達するとシナプス小胞が
細胞膜に融合しシナプス間隙に神経伝達物質が放出される。
そして拡散した神経伝達物質がシナプス後細胞に存在する
受容体に結合することで刺激が伝達されて行く。
シナプスにおける典型的な情報伝達機序は以下のように進む。
1. 前シナプス細胞の軸索を活動電位が伝わり、末端にある膨らみであるシナプス小頭に到達する
2. 活動電位によりシナプス小頭の膜上に位置する電位依存性カルシウムイオンチャネルが開く。
3. するとカルシウムイオンがシナプス内に流入し、シナプス小胞が細胞膜に接して神経伝達物質が細胞外に開口放出される。
4. 神経伝達物質はシナプス間隙を拡散し、後シナプス細胞の細胞膜上に分布する神経伝達物質受容体に結合する。
5. 後シナプス細胞のイオンチャネルが開き、細胞膜内外の電位差が変化する。
http://ja.wikipedia.org/wiki/%E3%82%B7%E3%83%8A%E3%83%97%E3%82%B9
参考文献:「脳のしくみがわかる本」寺沢宏次監修 P60
「シナプスは、1個の神経細胞に100個から10万個もある」
http://books.yahoo.co.jp/book_detail/31915842
ひとつのニューロンには、最大で数千の入出力(シナプス)が存在します。
http://plaza.rakuten.co.jp/neuron/diary/200609170000/
神経伝達物質はシナプス間隙に放出されると、拡散によって広がり、
後シナプス細胞の細胞膜上にある受容体と結びついて活性化される。
受容体がイオンチャネル型の場合そのイオンチャネルが開き、
受容体が代謝型であればその後いくつかのステップを経てイオンチャネルを開かせ、
後シナプス細胞に脱分極ないし過分極を生じさせる。
放出後は速やかに酵素によって不活性化されるか、
または前シナプス終末に再吸収され、一部は再びシナプス小胞に貯蔵され再利用される
(元のシナプス小胞に戻るのではなく別のシナプス小胞に充填される)。
http://ja.wikipedia.org/wiki/%E7%A5%9E%E7%B5%8C%E4%BC%9D%E9%81%94%E7%89%A9%E8%B3%AA
ナトリウムイオンが流入してきた部分は、イオン分布が変わってしまいます。
このままでは二度と興奮できないので、イオン分布はもとに戻ります。
(そうして膜電位ももとに戻ります。)
もとに戻る為には、ナトリウムイオンを細胞の外にくみ出して、
カリウムイオンを細胞の中にくみ入れなくてはなりません。
それを行っているのが、ナトリウムポンプと呼ばれるタンパク質です。
ナトリウムポンプは、ATPのエネルギ-を使い
能動輸送でナトリウムイオンを細胞の外にくみ出して、
カリウムイオンを細胞の中にくみ入れるのです。
http://www.fujijuku.net/biollec/16.HTML
ひとつの細胞に数千から数万に及ぶシナプス入力があります。
図6で模式的に単純化して説明します。
Aのように少数のシナプスへの興奮性入力ではたりませんが、
Bのようにそのようなシナプス入力がたくさん同時におこると、
それらが加算的に統合され、軸索の根元のあたりの膜電位が充分に浅くなると、
そこに存在する膜電位感受性ナトリウムチャネルが開いて、そこで活動電位が生じます。
http://www.tmin.ac.jp/medical/17/synapse2.html
活動電位からシナプス電位へ
シナプスにおいて、活動電位からシナプス電位に変換されます(図5)。
活動電位は、神経細胞の軸索突起の根元で発生し、
軸索をつたわってシナプス前部に達します。
シナプス前部には膜電位感受性カルシウムチャネルがありますので、
これが活動電位によって膜電位が上昇したことを感知して、活性化し、
つまりチャネルが開いてカルシウムが細胞内にはいります。
このカルシウムによって、シナプス前部にあったシナプス小胞が細胞膜に融合し、
中に入っていた神経伝達物質をシナプス間隙に放出します。
興奮性の神経伝達を司っているグルタミン酸を例にあげてみますと、
グルタミン酸がシナプス後膜にあるその受容体、
つまりグルタミン酸受容体に結合します。
グルタミン酸受容体は4つの膜蛋白からなり、
それらのまん中に穴(チャネル)を形成していて、
受容体にグルタミン酸が結合するとタンパク質の構造が変化して、短時間穴が開きます。
この穴はカチオン(陽イオン;ナトリウム、カリウム、カルシウムなど)
をとおす性質があるため、
細胞外には内側に比べて高濃度のナトリウムイオンがあり、
それがチャネルをとおって細胞内に流入します。
ナトリウムイオンはプラスですから、その結果、
細胞内のマイナスの程度が浅くなる、つまり脱分極します。
脱分極するにつれ、カリウムイオンの流出もあるために、
活動電位のように膜電位がプラスまで上昇することはありません。
このような神経伝達物質によって引き起された
シナプス後部の膜電位をシナプス電位といいます。
グルタミン酸 (Glu) の場合、シナプス電位は脱分極性なので
興奮性シナプス後電位といいます。
一方、GABA (ギャバ) やグリシンは抑制性です。
抑制性ニューロンからGABAが分泌されると、
GABA受容体チャネルが活性化し、塩素イオンを選択的に通す穴がひらき、
細胞外に多い塩素イオンが細胞内に流入します。塩素イオンはマイナスなので、
その結果膜電位はよりマイナスの方向に深くなり、つまり過分極になり、
これを抑制性シナプス後電位といいます。これによって、活動電位が出にくくなります(図5)。
麻酔薬、睡眠薬、抗不安薬などはこのGABA受容体の働きを促進させ、
グルタミン酸による興奮性の神経活動を抑制させるのです。
http://www.tmin.ac.jp/medical/17/synapse2.html
神経伝達と神経修飾
http://www.biol.s.u-tokyo.ac.jp/users/naibunpi/Oka/2005Summer_Seminar.pdf
3、興奮性シナプス後電位(EPSP) と、抑制性シナプス後電位(IPSP)
化学信号には、シナプスを興奮させる
(シナプス電位にプラス電位を発生させる)
ものと、シナプスを抑制する
(シナプス電位にプラス電位を発生させるのを抑制する)
ものの2種類がある
シナプスを興奮させる電位を興奮性シナプス後電位(EPSP)という
シナプスを抑制させる電位を抑制性シナプス後電位(IPSP)という
なお、神経細胞のシナプスにおいては
Na+-K+チャネルにより細胞内に興奮性シナプス後電位(EPSP)を発生させる
Cl-チャネルにより細胞内に抑制性シナプス後電位(IPSP)を発生させる
シナプス電位の電気信号は違う
以上のようなアナログ電気信号が
一つの化学信号によってシナプスに発生する電気信号であり、
複数の化学信号が入力されると、上記の電気信号が重ね合わさったような
アナログ電気信号となる
***参考資料***
参考文献:「生体電気信号とはなにか」 杉晴夫
活動電位とシナプス電位
脳の活動は活動電位とシナプス電位によって成り立っています。
活動電位は細胞体からシナプス前部まで軸索突起(アクソン)を伝わります。
活動電位が軸索の終末にくるとシナプス前部から神経伝達物質が分泌され、
それが樹状突起(デンドライト)のシナプス後部の受容体を活性化し、
シナプス後細胞にシナプス電位を発生させます。
神経細胞は数千から数万ものシナプス入力をうけており、
http://www.tmin.ac.jp/medical/17/synapse2.html
情報伝達は一方向に行われ、興奮がシナプスに達するとシナプス小胞が
細胞膜に融合しシナプス間隙に神経伝達物質が放出される。
そして拡散した神経伝達物質がシナプス後細胞に存在する
受容体に結合することで刺激が伝達されて行く。
シナプスにおける典型的な情報伝達機序は以下のように進む。
1. 前シナプス細胞の軸索を活動電位が伝わり、末端にある膨らみであるシナプス小頭に到達する
2. 活動電位によりシナプス小頭の膜上に位置する電位依存性カルシウムイオンチャネルが開く。
3. するとカルシウムイオンがシナプス内に流入し、シナプス小胞が細胞膜に接して神経伝達物質が細胞外に開口放出される。
4. 神経伝達物質はシナプス間隙を拡散し、後シナプス細胞の細胞膜上に分布する神経伝達物質受容体に結合する。
5. 後シナプス細胞のイオンチャネルが開き、細胞膜内外の電位差が変化する。
http://ja.wikipedia.org/wiki/%E3%82%B7%E3%83%8A%E3%83%97%E3%82%B9
参考文献:「脳のしくみがわかる本」寺沢宏次監修 P60
「シナプスは、1個の神経細胞に100個から10万個もある」
http://books.yahoo.co.jp/book_detail/31915842
ひとつのニューロンには、最大で数千の入出力(シナプス)が存在します。
http://plaza.rakuten.co.jp/neuron/diary/200609170000/
神経伝達物質はシナプス間隙に放出されると、拡散によって広がり、
後シナプス細胞の細胞膜上にある受容体と結びついて活性化される。
受容体がイオンチャネル型の場合そのイオンチャネルが開き、
受容体が代謝型であればその後いくつかのステップを経てイオンチャネルを開かせ、
後シナプス細胞に脱分極ないし過分極を生じさせる。
放出後は速やかに酵素によって不活性化されるか、
または前シナプス終末に再吸収され、一部は再びシナプス小胞に貯蔵され再利用される
(元のシナプス小胞に戻るのではなく別のシナプス小胞に充填される)。
http://ja.wikipedia.org/wiki/%E7%A5%9E%E7%B5%8C%E4%BC%9D%E9%81%94%E7%89%A9%E8%B3%AA
ナトリウムイオンが流入してきた部分は、イオン分布が変わってしまいます。
このままでは二度と興奮できないので、イオン分布はもとに戻ります。
(そうして膜電位ももとに戻ります。)
もとに戻る為には、ナトリウムイオンを細胞の外にくみ出して、
カリウムイオンを細胞の中にくみ入れなくてはなりません。
それを行っているのが、ナトリウムポンプと呼ばれるタンパク質です。
ナトリウムポンプは、ATPのエネルギ-を使い
能動輸送でナトリウムイオンを細胞の外にくみ出して、
カリウムイオンを細胞の中にくみ入れるのです。
http://www.fujijuku.net/biollec/16.HTML
ひとつの細胞に数千から数万に及ぶシナプス入力があります。
図6で模式的に単純化して説明します。
Aのように少数のシナプスへの興奮性入力ではたりませんが、
Bのようにそのようなシナプス入力がたくさん同時におこると、
それらが加算的に統合され、軸索の根元のあたりの膜電位が充分に浅くなると、
そこに存在する膜電位感受性ナトリウムチャネルが開いて、そこで活動電位が生じます。
http://www.tmin.ac.jp/medical/17/synapse2.html
活動電位からシナプス電位へ
シナプスにおいて、活動電位からシナプス電位に変換されます(図5)。
活動電位は、神経細胞の軸索突起の根元で発生し、
軸索をつたわってシナプス前部に達します。
シナプス前部には膜電位感受性カルシウムチャネルがありますので、
これが活動電位によって膜電位が上昇したことを感知して、活性化し、
つまりチャネルが開いてカルシウムが細胞内にはいります。
このカルシウムによって、シナプス前部にあったシナプス小胞が細胞膜に融合し、
中に入っていた神経伝達物質をシナプス間隙に放出します。
興奮性の神経伝達を司っているグルタミン酸を例にあげてみますと、
グルタミン酸がシナプス後膜にあるその受容体、
つまりグルタミン酸受容体に結合します。
グルタミン酸受容体は4つの膜蛋白からなり、
それらのまん中に穴(チャネル)を形成していて、
受容体にグルタミン酸が結合するとタンパク質の構造が変化して、短時間穴が開きます。
この穴はカチオン(陽イオン;ナトリウム、カリウム、カルシウムなど)
をとおす性質があるため、
細胞外には内側に比べて高濃度のナトリウムイオンがあり、
それがチャネルをとおって細胞内に流入します。
ナトリウムイオンはプラスですから、その結果、
細胞内のマイナスの程度が浅くなる、つまり脱分極します。
脱分極するにつれ、カリウムイオンの流出もあるために、
活動電位のように膜電位がプラスまで上昇することはありません。
このような神経伝達物質によって引き起された
シナプス後部の膜電位をシナプス電位といいます。
グルタミン酸 (Glu) の場合、シナプス電位は脱分極性なので
興奮性シナプス後電位といいます。
一方、GABA (ギャバ) やグリシンは抑制性です。
抑制性ニューロンからGABAが分泌されると、
GABA受容体チャネルが活性化し、塩素イオンを選択的に通す穴がひらき、
細胞外に多い塩素イオンが細胞内に流入します。塩素イオンはマイナスなので、
その結果膜電位はよりマイナスの方向に深くなり、つまり過分極になり、
これを抑制性シナプス後電位といいます。これによって、活動電位が出にくくなります(図5)。
麻酔薬、睡眠薬、抗不安薬などはこのGABA受容体の働きを促進させ、
グルタミン酸による興奮性の神経活動を抑制させるのです。
http://www.tmin.ac.jp/medical/17/synapse2.html
神経伝達と神経修飾
http://www.biol.s.u-tokyo.ac.jp/users/naibunpi/Oka/2005Summer_Seminar.pdf
3、興奮性シナプス後電位(EPSP) と、抑制性シナプス後電位(IPSP)
化学信号には、シナプスを興奮させる
(シナプス電位にプラス電位を発生させる)
ものと、シナプスを抑制する
(シナプス電位にプラス電位を発生させるのを抑制する)
ものの2種類がある
シナプスを興奮させる電位を興奮性シナプス後電位(EPSP)という
シナプスを抑制させる電位を抑制性シナプス後電位(IPSP)という
なお、神経細胞のシナプスにおいては
Na+-K+チャネルにより細胞内に興奮性シナプス後電位(EPSP)を発生させる
Cl-チャネルにより細胞内に抑制性シナプス後電位(IPSP)を発生させる

(1)Na+-K+チャネルにより発生する興奮性シナプス後電位(EPSP)
①拡散電流
拡散電流 = Na+拡散電流(拡散電位:-60mVによる細胞外から細胞内への電流)
+ K+拡散電流(拡散電位:90mVによる細胞内から細胞外への電流)
= 15mVの拡散電流
15mVの拡散電流となるのは、
Na+とK+に対するイオンチャネルの抵抗の関係でこうなる
(この拡散電流の計算は、別の記事で計算します)
つまり、拡散電流はK+拡散電流成分の方が大きく、
細胞外への流出する方に流れることになる
②電圧電流
電圧電流 = Na+電圧電流(膜電位の電圧電流)
③膜を流れる電流
膜を流れる電流=拡散電流+電圧電流=15mV+膜電位の電流
となり、イオンチャネルがいくら開いても
電圧電流が拡散電流(15mV)と平衡する
-15mVの膜電位までしかならない
(2)Cl-チャネルにより発生する抑制性シナプス後電位(IPSP)
①拡散電流
拡散電流 = Cl-拡散電流(拡散電位:70mVによる電流)
②電圧電流
電圧電流 = Cl-電圧電流(膜電位による電圧電流)
③膜を流れる電流
膜を流れる電流=拡散電流+電圧電流=70mV+膜電位による電流
となり、Cl-チャネルは、静止膜電位(-70mV)と同じ電位にするように作用する
つまり、プラス電位を発生させるのを抑制するような働きがある
なお、細胞によってCl-の平衡電位が違う場合は、
Cl-の平衡電位によって以下のように変動する
Cl-の平衡電位が静止膜電位より低い場合は、静止膜電位より
マイナス方向の電位を発生させる
Cl-の平衡電位が静止膜電位より高い場合は、静止膜電位より
プラス方向の電位を発生させる
このようにCl-の平衡電位によって、
静止膜電位よりプラス方向の電位を発生させるのか
マイナス方向の電位を発生させるのかが決まるので、
必ずマイナス電位を発生させるなどとは一概にはいえない
実際にCl-チャネルによりプラス電位を発生させる例もある
***参考資料***
神経細胞から神経細胞へシナプスを介してシグナルが伝達されますが、
シナプスには興奮を伝達する興奮性シナプスだけではなく、
興奮を抑制する抑制性シナプスもあります。
興奮性シナプスはグルタミン酸などの興奮性神経伝達物質を含み、
この神経伝達物質がシナプス後膜に達すると脱分極(Depolarization)を起こし、
興奮性シナプス後電位(EPSP)が発生します。
これはナトリウムやカルシウムイオンチャンネルが開き、
イオンが流入することにより起こります。
GABAやグリシンなどを含むシナプスは抑制性シナプスです。
これらの神経伝達物質の放出によってマイナスイオンである塩素イオンが流入し、
過分極(Hyperpolarization)が起こり抑制性シナプス後電位(IPSP)が発生します。
http://bunseiri.hp.infoseek.co.jp/Sinkeihp.htm
興奮性の神経伝達を司っているグルタミン酸を例にあげてみますと、
グルタミン酸がシナプス後膜にあるその受容体、
つまりグルタミン酸受容体に結合します。
グルタミン酸受容体は4つの膜蛋白からなり、
それらのまん中に穴(チャネル)を形成していて、
受容体にグルタミン酸が結合するとタンパク質の構造が変化して、短時間穴が開きます。
この穴はカチオン(陽イオン;ナトリウム、カリウム、カルシウムなど)をとおす性質があるため、
細胞外には内側に比べて高濃度のナトリウムイオンがあり、
それがチャネルをとおって細胞内に流入します。
ナトリウムイオンはプラスですから、その結果、
細胞内のマイナスの程度が浅くなる、つまり脱分極します。
脱分極するにつれ、カリウムイオンの流出もあるために、
活動電位のように膜電位がプラスまで上昇することはありません。
このような神経伝達物質によって引き起されたシナプス後部の膜電位をシナプス電位といいます。
グルタミン酸 (Glu) の場合、シナプス電位は脱分極性なので興奮性シナプス後電位といいます。
一方、GABA (ギャバ) やグリシンは抑制性です。抑制性ニューロンからGABAが分泌されると、
GABA受容体チャネルが活性化し、塩素イオンを選択的に通す穴がひらき、
細胞外に多い塩素イオンが細胞内に流入します。塩素イオンはマイナスなので、
その結果膜電位はよりマイナスの方向に深くなり、
つまり過分極になり、これを抑制性シナプス後電位といいます。
http://www.tmin.ac.jp/medical/17/synapse2.html
4、活動電位
(1)活動電位の説明
化学シナプスにより、ある一定の量以上のシナプスを興奮させる化学信号が到達すると
受容体で化学信号を受け取り、Na+とK+イオンを通すNa+-K+イオンチャネルが開くことにより
Na+とK+イオン電流がたくさん流れて多数の興奮性シナプス後電位(EPSP)が発生し、
シナプス電位が次第に上昇していく。
あるいは電気シナプスにより、他のシナプスとの電位差に応じたイオン電流が流れて
シナプス電位が次第に上昇していく。
そしてシナプス電位がある値(しきい値、閾値)を越えると、
化学物質によりチャネルが開くリガンド依存性チャネルとは別のチャネルで、
シナプス電位によって開く電位依存性チャネルのNa+チャネルとK+チャネルが開
①拡散電流
拡散電流 = Na+拡散電流(拡散電位:-60mVによる細胞外から細胞内への電流)
+ K+拡散電流(拡散電位:90mVによる細胞内から細胞外への電流)
= 15mVの拡散電流
15mVの拡散電流となるのは、
Na+とK+に対するイオンチャネルの抵抗の関係でこうなる
(この拡散電流の計算は、別の記事で計算します)
つまり、拡散電流はK+拡散電流成分の方が大きく、
細胞外への流出する方に流れることになる
②電圧電流
電圧電流 = Na+電圧電流(膜電位の電圧電流)
③膜を流れる電流
膜を流れる電流=拡散電流+電圧電流=15mV+膜電位の電流
となり、イオンチャネルがいくら開いても
電圧電流が拡散電流(15mV)と平衡する
-15mVの膜電位までしかならない
(2)Cl-チャネルにより発生する抑制性シナプス後電位(IPSP)
①拡散電流
拡散電流 = Cl-拡散電流(拡散電位:70mVによる電流)
②電圧電流
電圧電流 = Cl-電圧電流(膜電位による電圧電流)
③膜を流れる電流
膜を流れる電流=拡散電流+電圧電流=70mV+膜電位による電流
となり、Cl-チャネルは、静止膜電位(-70mV)と同じ電位にするように作用する
つまり、プラス電位を発生させるのを抑制するような働きがある
なお、細胞によってCl-の平衡電位が違う場合は、
Cl-の平衡電位によって以下のように変動する
Cl-の平衡電位が静止膜電位より低い場合は、静止膜電位より
マイナス方向の電位を発生させる
Cl-の平衡電位が静止膜電位より高い場合は、静止膜電位より
プラス方向の電位を発生させる
このようにCl-の平衡電位によって、
静止膜電位よりプラス方向の電位を発生させるのか
マイナス方向の電位を発生させるのかが決まるので、
必ずマイナス電位を発生させるなどとは一概にはいえない
実際にCl-チャネルによりプラス電位を発生させる例もある
***参考資料***
神経細胞から神経細胞へシナプスを介してシグナルが伝達されますが、
シナプスには興奮を伝達する興奮性シナプスだけではなく、
興奮を抑制する抑制性シナプスもあります。
興奮性シナプスはグルタミン酸などの興奮性神経伝達物質を含み、
この神経伝達物質がシナプス後膜に達すると脱分極(Depolarization)を起こし、
興奮性シナプス後電位(EPSP)が発生します。
これはナトリウムやカルシウムイオンチャンネルが開き、
イオンが流入することにより起こります。
GABAやグリシンなどを含むシナプスは抑制性シナプスです。
これらの神経伝達物質の放出によってマイナスイオンである塩素イオンが流入し、
過分極(Hyperpolarization)が起こり抑制性シナプス後電位(IPSP)が発生します。
http://bunseiri.hp.infoseek.co.jp/Sinkeihp.htm
興奮性の神経伝達を司っているグルタミン酸を例にあげてみますと、
グルタミン酸がシナプス後膜にあるその受容体、
つまりグルタミン酸受容体に結合します。
グルタミン酸受容体は4つの膜蛋白からなり、
それらのまん中に穴(チャネル)を形成していて、
受容体にグルタミン酸が結合するとタンパク質の構造が変化して、短時間穴が開きます。
この穴はカチオン(陽イオン;ナトリウム、カリウム、カルシウムなど)をとおす性質があるため、
細胞外には内側に比べて高濃度のナトリウムイオンがあり、
それがチャネルをとおって細胞内に流入します。
ナトリウムイオンはプラスですから、その結果、
細胞内のマイナスの程度が浅くなる、つまり脱分極します。
脱分極するにつれ、カリウムイオンの流出もあるために、
活動電位のように膜電位がプラスまで上昇することはありません。
このような神経伝達物質によって引き起されたシナプス後部の膜電位をシナプス電位といいます。
グルタミン酸 (Glu) の場合、シナプス電位は脱分極性なので興奮性シナプス後電位といいます。
一方、GABA (ギャバ) やグリシンは抑制性です。抑制性ニューロンからGABAが分泌されると、
GABA受容体チャネルが活性化し、塩素イオンを選択的に通す穴がひらき、
細胞外に多い塩素イオンが細胞内に流入します。塩素イオンはマイナスなので、
その結果膜電位はよりマイナスの方向に深くなり、
つまり過分極になり、これを抑制性シナプス後電位といいます。
http://www.tmin.ac.jp/medical/17/synapse2.html
4、活動電位
(1)活動電位の説明
化学シナプスにより、ある一定の量以上のシナプスを興奮させる化学信号が到達すると
受容体で化学信号を受け取り、Na+とK+イオンを通すNa+-K+イオンチャネルが開くことにより
Na+とK+イオン電流がたくさん流れて多数の興奮性シナプス後電位(EPSP)が発生し、
シナプス電位が次第に上昇していく。
あるいは電気シナプスにより、他のシナプスとの電位差に応じたイオン電流が流れて
シナプス電位が次第に上昇していく。
そしてシナプス電位がある値(しきい値、閾値)を越えると、
化学物質によりチャネルが開くリガンド依存性チャネルとは別のチャネルで、
シナプス電位によって開く電位依存性チャネルのNa+チャネルとK+チャネルが開
ただし、K+チャネルはすぐには開ききらないでだんだんと開くので
しきい値電圧に到達した時間から約0.5msぐらいの間は、
Na+チャネルによるNa+のイオン電流の方が多い
それで、細胞内へと流れるNa+のイオン電流は、
膜を流れる電流
= Na+の拡散電位約-60mVによる拡散電流 + Na+のシナプス電位による電圧電流
の電流が流れて一気にNa+の平衡電位である60mV近くまでシナプス電位が上昇する
実際は、K+チャネルも多少開いているので細胞外へと流れるK+イオン電流による下落があり
約30mV程度まで上昇する
そしてその後は、
電位依存性チャネルのNa+チャネルが閉じ始めて
細胞内へと流れるNa+イオン電流がだんだんと小さくなる一方で、
電位依存性チャネルのK+チャネルが完全に開くので
Na+イオン電流よりもK+チャネルによる細胞外へと流れるK+イオン電流の方が大きくなる
細胞外へと流れるK+のイオン電流は、
膜を流れる電流
=K+の拡散電位約90mVによる拡散電流 +
K+のシナプス電位による電圧電流
=約120mVによる電流
となり、急激にシナプス電位がマイナス電位へと下落する
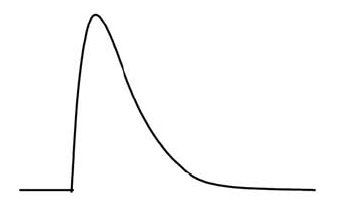
このようにしてシナプス電位が1ミリ秒以下の短時間だけ
急激にプラス電位(最大で約30mV)になる現象が発生します。
この1ミリ秒以下の短時間だけプラス電位になる電圧波形の信号を活動電位という
そしてこの活動電位が、神経細胞(ニューロン)の軸索を伝達する
しきい値電圧に到達した時間から約0.5msぐらいの間は、
Na+チャネルによるNa+のイオン電流の方が多い
それで、細胞内へと流れるNa+のイオン電流は、
膜を流れる電流
= Na+の拡散電位約-60mVによる拡散電流 + Na+のシナプス電位による電圧電流
の電流が流れて一気にNa+の平衡電位である60mV近くまでシナプス電位が上昇する
実際は、K+チャネルも多少開いているので細胞外へと流れるK+イオン電流による下落があり
約30mV程度まで上昇する
そしてその後は、
電位依存性チャネルのNa+チャネルが閉じ始めて
細胞内へと流れるNa+イオン電流がだんだんと小さくなる一方で、
電位依存性チャネルのK+チャネルが完全に開くので
Na+イオン電流よりもK+チャネルによる細胞外へと流れるK+イオン電流の方が大きくなる
細胞外へと流れるK+のイオン電流は、
膜を流れる電流
=K+の拡散電位約90mVによる拡散電流 +
K+のシナプス電位による電圧電流
=約120mVによる電流
となり、急激にシナプス電位がマイナス電位へと下落する
このようにしてシナプス電位が1ミリ秒以下の短時間だけ
急激にプラス電位(最大で約30mV)になる現象が発生します。
この1ミリ秒以下の短時間だけプラス電位になる電圧波形の信号を活動電位という
そしてこの活動電位が、神経細胞(ニューロン)の軸索を伝達する
膜電位がプラスに反転する現象を脱分極という
なお閾値をこえると、化学信号の入力があってもなくても
自動的に一定の振幅(電位幅)、一定の時間幅の活動電位を発生させる
つまり活動電位のパルス電気信号が発生している1ミリ秒の間は、
(実際はもう1ミリ秒足した2ミリ秒の間
詳細は、絶対不応期の項で解説)
化学信号の入力を受けつけていないということであり、
抑制性シナプス後電位を発生させる化学信号がきていても
それによって活動電位の電位が下がることはないし、
興奮性シナプス後電位を発生させる化学信号がきていても
それによって活動電位の電位が上がることはない
このように、活動電位はアナログ的に様々な波形を描くことがなく
閾値を超えたら一定の振幅(電位幅)、一定の時間幅の活動電位を発生させて、
閾値を超えなかったら活動電位を発生させないという性質がある
活動電位が、発生するかしないかの2つの状態しか取らないことを
全か無かの法則(all-or-none law)という
したがって、シナプス電位が様々な振幅値を取るアナログ信号なのに対して
活動電位は発生するかしないかの2つの状態しかないので
ディジタル信号だと考えることができる
なお、この活動電位は、ノーベル賞化学者のホジキンとハクスレーにより
完璧に説明する回路モデルが提唱され、
4次元の非線形微分方程式を解くことにより活動電位の波形を求めることができる
通称、HHモデルといわれるHodgkin-Huxley モデルは、
正確な活動電位の発生を表現するものとして世界で最も有名なモデルであり
絶大な信頼を得ている
参照:Hodgkin-Huxley モデル
http://kirigakure.dyndns.org/~brain/pukiwiki-1.4.5_1/index.php?Hodgkin-Huxley
http://www.cns.atr.jp/~doya/naist/mathsci/icns02.pdf
ただし、Hodgkin-Huxley モデルは、活動電位の信号波形を正確に説明できるが
活動電位が発生する前のシナプス電位の変化は
説明することはできないことに注意する必要がある
あくまでHodgkin-Huxley モデルは、活動電位の信号波形を正確に説明するモデルである
(2)活動電位発生の簡単な数式表現(McCulloch-Pittsモデル)
以上の情報を、Hodgkin-Huxley モデルによる
4次元の非線形微分方程式のような難しい数式ではなく
簡単な数式で、活動電位が発生する前のシナプス電位の変化も含めて説明すると
以下のようになる
シナプス電位:P (アナログ電気信号)
興奮性シナプス後電位:EPSP
抑制性シナプス後電位:IPSP
P = { EPSP1+EPSP2+・・・+EPSPn} + { IPSP1+IPSP2+・・・+IPSPm }
n、m:数千から数万
P >= 活動電位が発生する閾値電圧
のとき一定の振幅、一定の時間幅を持った活動電位(電気パルス信号)が発生する
参照:ニューロンモデルの一つであるMcCulloch-Pittsモデル
http://www.cns.atr.jp/~doya/naist/mathsci/icns02.pdf
なお、この数式モデルは非常に簡易なモデルであり、
実際のシナプス電位を正確に表現できていないことに注意する必要がある
厳密に表現しようとすればやはり、Hodgkin-Huxley モデルのような
等価回路モデルによって表現するしかない
(3)活動電位の電圧波形
電圧幅:約100mV
時間幅:約1ms
周波数成分:約1kHz~約数十kHz
の電気パルス信号=ディジタル信号である
周波数1kの根拠 :f=1/1ms=1k
周波数数十kの根拠:約1msの時間幅があるパルスを
フーリエ解析するとさらに高周波成分があると推測されるから
実際に筋電位で計測される周波数の最大値は、数10KHz~数100kHz
***参考資料***
筋電位の最小単位である運動単位の活動電位は
ほんの数ミリ秒しか持続しないパルスのような電位であるが、
表面電極で検出される表面筋電位はそれが無数に加算されたものなので、
連続的に発生する雑音のような電位となっている。
従って、様々な電位・周波数・位相の交流信号が時間的にも
変化しながら加算されていると見なせる。
その平均電位は数μV~数mV程度、周波数成分は数Hz~数100kHz程度である。
極で拾い上げられた筋電位は数μV~数mV程度であり、
これを通常の記録・保存装置に入力できる電位であるVオーダまで増幅するのには、
1,000~10,000倍に増幅する必要がある。アンプは、入力された信号を忠実に増幅する、
つまり入力信号と出力信号が全く相似形になるように増幅することが要求される。
そのための要件とは、十分な出力振幅幅(ダイナミックレンジ)と
高い対ノイズ比(S/N)を持っていること、
十分広い周波数帯域を持っていること、
周波数成分間の位相変位が十分小さいことが要求される。
表面筋電位を記録するアンプとしては、
ダイナミックレンジは 48dB、S/Nは30dB
(ただし増幅率を必要に応じて変化させる場合はこれより低くて良い)、
周波数成分は数Hz~数10KHz程度である。
http://homepage2.nifty.com/aseo/emg.htm
(4)活動電位の速度
活動電位は、太さ以外の条件が同じであるとき、より太い軸索上でより速く伝導する。
その速さは1-1000m/sである。
なお閾値をこえると、化学信号の入力があってもなくても
自動的に一定の振幅(電位幅)、一定の時間幅の活動電位を発生させる
つまり活動電位のパルス電気信号が発生している1ミリ秒の間は、
(実際はもう1ミリ秒足した2ミリ秒の間
詳細は、絶対不応期の項で解説)
化学信号の入力を受けつけていないということであり、
抑制性シナプス後電位を発生させる化学信号がきていても
それによって活動電位の電位が下がることはないし、
興奮性シナプス後電位を発生させる化学信号がきていても
それによって活動電位の電位が上がることはない
このように、活動電位はアナログ的に様々な波形を描くことがなく
閾値を超えたら一定の振幅(電位幅)、一定の時間幅の活動電位を発生させて、
閾値を超えなかったら活動電位を発生させないという性質がある
活動電位が、発生するかしないかの2つの状態しか取らないことを
全か無かの法則(all-or-none law)という
したがって、シナプス電位が様々な振幅値を取るアナログ信号なのに対して
活動電位は発生するかしないかの2つの状態しかないので
ディジタル信号だと考えることができる
なお、この活動電位は、ノーベル賞化学者のホジキンとハクスレーにより
完璧に説明する回路モデルが提唱され、
4次元の非線形微分方程式を解くことにより活動電位の波形を求めることができる
通称、HHモデルといわれるHodgkin-Huxley モデルは、
正確な活動電位の発生を表現するものとして世界で最も有名なモデルであり
絶大な信頼を得ている
参照:Hodgkin-Huxley モデル
http://kirigakure.dyndns.org/~brain/pukiwiki-1.4.5_1/index.php?Hodgkin-Huxley
http://www.cns.atr.jp/~doya/naist/mathsci/icns02.pdf
ただし、Hodgkin-Huxley モデルは、活動電位の信号波形を正確に説明できるが
活動電位が発生する前のシナプス電位の変化は
説明することはできないことに注意する必要がある
あくまでHodgkin-Huxley モデルは、活動電位の信号波形を正確に説明するモデルである
(2)活動電位発生の簡単な数式表現(McCulloch-Pittsモデル)
以上の情報を、Hodgkin-Huxley モデルによる
4次元の非線形微分方程式のような難しい数式ではなく
簡単な数式で、活動電位が発生する前のシナプス電位の変化も含めて説明すると
以下のようになる
シナプス電位:P (アナログ電気信号)
興奮性シナプス後電位:EPSP
抑制性シナプス後電位:IPSP
P = { EPSP1+EPSP2+・・・+EPSPn} + { IPSP1+IPSP2+・・・+IPSPm }
n、m:数千から数万
P >= 活動電位が発生する閾値電圧
のとき一定の振幅、一定の時間幅を持った活動電位(電気パルス信号)が発生する
参照:ニューロンモデルの一つであるMcCulloch-Pittsモデル
http://www.cns.atr.jp/~doya/naist/mathsci/icns02.pdf
なお、この数式モデルは非常に簡易なモデルであり、
実際のシナプス電位を正確に表現できていないことに注意する必要がある
厳密に表現しようとすればやはり、Hodgkin-Huxley モデルのような
等価回路モデルによって表現するしかない
(3)活動電位の電圧波形
電圧幅:約100mV
時間幅:約1ms
周波数成分:約1kHz~約数十kHz
の電気パルス信号=ディジタル信号である
周波数1kの根拠 :f=1/1ms=1k
周波数数十kの根拠:約1msの時間幅があるパルスを
フーリエ解析するとさらに高周波成分があると推測されるから
実際に筋電位で計測される周波数の最大値は、数10KHz~数100kHz
***参考資料***
筋電位の最小単位である運動単位の活動電位は
ほんの数ミリ秒しか持続しないパルスのような電位であるが、
表面電極で検出される表面筋電位はそれが無数に加算されたものなので、
連続的に発生する雑音のような電位となっている。
従って、様々な電位・周波数・位相の交流信号が時間的にも
変化しながら加算されていると見なせる。
その平均電位は数μV~数mV程度、周波数成分は数Hz~数100kHz程度である。
極で拾い上げられた筋電位は数μV~数mV程度であり、
これを通常の記録・保存装置に入力できる電位であるVオーダまで増幅するのには、
1,000~10,000倍に増幅する必要がある。アンプは、入力された信号を忠実に増幅する、
つまり入力信号と出力信号が全く相似形になるように増幅することが要求される。
そのための要件とは、十分な出力振幅幅(ダイナミックレンジ)と
高い対ノイズ比(S/N)を持っていること、
十分広い周波数帯域を持っていること、
周波数成分間の位相変位が十分小さいことが要求される。
表面筋電位を記録するアンプとしては、
ダイナミックレンジは 48dB、S/Nは30dB
(ただし増幅率を必要に応じて変化させる場合はこれより低くて良い)、
周波数成分は数Hz~数10KHz程度である。
http://homepage2.nifty.com/aseo/emg.htm
(4)活動電位の速度
活動電位は、太さ以外の条件が同じであるとき、より太い軸索上でより速く伝導する。
その速さは1-1000m/sである。

***参考資料***
ホジキンとハクスレーはNa説を立てた(1945年)。軸索内部はK+濃度が高く,外
液ではNa+とCl-濃度が高い。静止状態では膜はK+に対して透過性が高く,Na+
に対しては低い。するとK+は外液の方へ流出する。その結果,膜の内側は外側に
対して負になる。負の電位はK+を引き付け,K+の外への移動をある程度制限し,
平衡となる。Na+も少し透過し,その総合的な結果が静止電位となる。
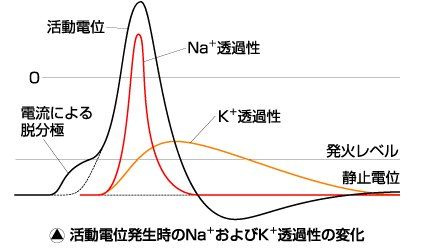
活動電位が発生すると,膜のNa+に対する透過性が選択的に高まり,Na+は内向
きに流入して膜電位を+側へ変化させる(脱分極)。その後,Na+の透過性が急激に
減少し,同時にK+の透過性が増加してくることによって後電位が現れる(図 活動
電位発生時のNa+およびK+透過性の変化)。
このような仕組みで活動電位が発生すると,そのたびに軸索は少量のNa+を受け
取り,同量のK+を失うことになる。そこでNa+を追い出し,K+を取り込んで,膜
の内外のイオン濃度を一定に保つようにしないと,軸索は働かなくなる。そこで
Na+の排出機序をNaポンプという。KポンプやClポンプもあるが,Naポンプが一
番重要な役割をしていると考えられる。
膜の興奮部では電位の逆転に伴い,表面から内部に向かう短時間の電流(局部電
流)が観察され,この電流によって隣接部が興奮する。この興奮によって隣接部の
膜の透過性が変化してNa+の移動が起こる。このようにして,電位の変動がつぎつ
ぎに神経を伝わっていくことになる。
透過性の実体は電位依存型Na+チャネル,電位依存型K+チャネルと呼
ばれるイオンチャネルであることが判明している。チャネルはポリペプチドである
(図 電位依存性Na+チャネルの分子構造)。チャネルは活性化ゲートと不活性化ゲ
ートをもち,静止時には活性化ゲートは閉じ,不活性化ゲートは開いている(次図
(イ))。脱分極が起こるとNa+が移動する(次図(ロ))。脱分極が続くと,活性化ゲー
トは開いているが,不活性化ゲートが閉じるためにイオンの移動は止まる(次図(ハ))。
再分極されると,不活性化ゲートが開き活性化ゲートが閉じて元の状態(次図(イ))
に戻る。
http://www.keirinkan.com/kori/kori_biology/kori_biology_n1/contents/bi-n1/4-bu/4-1-B.htm
活動電位(かつどうでんい、 action potential)は、なんらかの刺激に応じて
細胞膜に沿って流れる微弱な電位変化のこと。主としてナトリウムイオン、
カリウムイオンのイオンチャネルを通した受動的拡散でそれらイオンの細胞内外の
濃度差を変えることにより起きるものである。
細胞の内と外の間では、電位差が常に存在している。これは細胞内外でのイオン分布と、
これらイオンに対する細胞膜の透過性(特定のイオンの通しやすさ)に由来する。
電荷を持つイオンの分布が細胞内外で異なるため、活性化してない静止状態の
細胞の電位差は通常、細胞外と比べ細胞内がマイナスとなっている。
活動電位とは、この電位差がなんらかの刺激によって一時的に逆転する現象である。
活動電位はスパイクやインパルスともよばれる。また、活動電位に達することを
「発火する」(英 fire)と称することもある。
活動電位の速度と複雑さは細胞の種類により異なるものの、電位逆転の幅はだいたい同じである。
活動電位における負から正への電位の変化に要する時間は短く、数ミリ秒である。
どんな細胞の活動電位にも順に脱分極相、再分極相があり、多くの場合過分極相の段階もある。
活動電位は、太さ以外の条件が同じであるとき、より太い軸索上でより速く伝導する。
その速さは100-1000m/sである。
活動電位について知るためには、まず静止膜電位(静止電位ともいう)についておおまかに知る必要がある。
すべての細胞の膜内外に存在する電位差は通常、細胞外と比べ細胞内がマイナスである。
この状態を、膜は分極しているという。
活動していない状態にある膜の電位差は静止電位と呼ばれ、
神経細胞ではおよそ-70mVである(細胞外の電位を0としている)。
この電位差が生ずる要因はいくつかあるが、
もっとも重要なものは膜間における、イオン輸送と選択的なイオン透過性である。
活動電位の大まかな流れは次のようになっている。
[編集] 静止電位
静止電位において、いくつかのK+漏洩チャネルは開いている一方、
電位依存性Na+チャネルは閉じている。正味の電流は流れていないが、
膜間を移動している主なイオンはK+であり、その結果静止電位はK+平衡電位に比較近い値をとる。
[編集] 刺激
興奮刺激による膜の局所的な脱分極は、神経細胞の表面の膜にある電位依存性Na+チャネルを開く。
その結果Na+は濃度勾配および電気的勾配が推進力となり、細胞内へ流入する。
[編集] 脱分極
Na+が流入し膜電位の負電荷が減少するに従い、さらなるNa+チャネルが開き、
さらに大きなNa+の流入が引き起こされる。これは正のフィードバックのよい例である。
Na+チャネルが多く開くにつれ、Na+による電流はK+漏洩チャネルによる電流に打ち勝ち、
膜電位が逆転し内側がプラスとなる。
電位変化はNa+のみの移動によるものと考えるのではなく、
Na+とK+の膜間での「透過性の比」の変化によるものと考えるべきである。
この、脱分極とそれにともなうNa+チャネルの開口が周囲に広がっていくことで活動電位の伝導が起こる。
[編集] ピーク
膜電位が+30mV程度になると、Na+チャネルの電位感受性不活性化ゲートが閉じ、
さらなるNa+の流入を阻害する。それと同時に、
電位依存性K+チャネルの電位感受性活性化ゲートが開く。
活動電位のどの段階においても、きわめて少しのイオンの移動しか起こらないという
認識は重要である。活動電位が生じている間の、
細胞内外でのNa+とK+の濃度変化は無視できるほど極めて小さい。
http://ja.wikipedia.org/wiki/%E6%B4%BB%E5%8B%95%E9%9B%BB%E4%BD%8D
5、再分極
シナプス電位が+30mV程度になると、プラスイオンの流入が止まり、
逆にプラスイオンの流出が始まり急激にシナプス電位が元のマイナス電位に戻る
この再度マイナス電位に戻ることを再分極という
***参考資料***
再分極
電位依存性K+チャネルが開くことで、濃度勾配および電気的勾配が推進力となり
K+の流出が始まる。K+が流出することで、膜電位の逆転と再分極が引き起こされる。
http://ja.wikipedia.org/wiki/%E6%B4%BB%E5%8B%95%E9%9B%BB%E4%BD%8D
6、過分極
活動電位が立ち下がっているときはK+チャネルが完全に開いている状態なので、
K+の拡散電位:90mVにより、細胞内から細胞外へとK+イオン電流が流れる
K+の拡散電流は、
膜を通る電流 =
K+の90mVの拡散電位による拡散電流 + K+の膜電位による電圧電流=0
となる膜電位でとまることになり、
当然その膜電位は、-90mVとなる
このようにしてK+の平衡電位の-90mVになるまでシナプス電位が下がることになる
膜電位が通常の静止膜電位(-70mV)よりも低くなる状態を過分極という
再分極した後、一時的に過分極状態になる
他には、抑制性シナプス後電位によって過分極状態になることがある
***参考資料***
電位依存性K+チャネルの閉鎖は電位および時間に依存している。
このチャネルは膜電位の変化にすぐには応答せず、遅れて応答を返す。
そのため、膜が十分再分極した後もK+の流出が続き、一時的に膜電位が通常の静止電位より
もさらに低くなる。この過分極状態はアンダーシュートとも呼ばれる。
http://ja.wikipedia.org/wiki/%E6%B4%BB%E5%8B%95%E9%9B%BB%E4%BD%8D
7、不応期と活動電位の最大周波数
(1)絶対不応期と相対不応期
活動電位が発生した後、2ミリ秒程度は活動電位を発生できない。
この時期が絶対不応期という
(活動電位のパルスの立ち上がりから2ミリ秒であって、パルスの終了から2ミリ秒ではない
なので絶対不応期の2ミリ秒のうち1ミリ秒は活動電位のパルス波形の時間となる)
さらにその絶対不応期後の数ミリ秒は活動電位が発生しにくい。
この時期を相対不応期という
だいたい活動電位が発生した後の2ミリ秒から10ミリ秒ぐらい相対不応期がある
絶対不応期と相対不応期以上の期間では、
通常の大きな膜電位変化(約100mV)で活動電位を発生させることができるが、
相対不応期の活動電位の振幅は通常より小さくなる
ホジキンとハクスレーはNa説を立てた(1945年)。軸索内部はK+濃度が高く,外
液ではNa+とCl-濃度が高い。静止状態では膜はK+に対して透過性が高く,Na+
に対しては低い。するとK+は外液の方へ流出する。その結果,膜の内側は外側に
対して負になる。負の電位はK+を引き付け,K+の外への移動をある程度制限し,
平衡となる。Na+も少し透過し,その総合的な結果が静止電位となる。
活動電位が発生すると,膜のNa+に対する透過性が選択的に高まり,Na+は内向
きに流入して膜電位を+側へ変化させる(脱分極)。その後,Na+の透過性が急激に
減少し,同時にK+の透過性が増加してくることによって後電位が現れる(図 活動
電位発生時のNa+およびK+透過性の変化)。
このような仕組みで活動電位が発生すると,そのたびに軸索は少量のNa+を受け
取り,同量のK+を失うことになる。そこでNa+を追い出し,K+を取り込んで,膜
の内外のイオン濃度を一定に保つようにしないと,軸索は働かなくなる。そこで
Na+の排出機序をNaポンプという。KポンプやClポンプもあるが,Naポンプが一
番重要な役割をしていると考えられる。
膜の興奮部では電位の逆転に伴い,表面から内部に向かう短時間の電流(局部電
流)が観察され,この電流によって隣接部が興奮する。この興奮によって隣接部の
膜の透過性が変化してNa+の移動が起こる。このようにして,電位の変動がつぎつ
ぎに神経を伝わっていくことになる。
透過性の実体は電位依存型Na+チャネル,電位依存型K+チャネルと呼
ばれるイオンチャネルであることが判明している。チャネルはポリペプチドである
(図 電位依存性Na+チャネルの分子構造)。チャネルは活性化ゲートと不活性化ゲ
ートをもち,静止時には活性化ゲートは閉じ,不活性化ゲートは開いている(次図
(イ))。脱分極が起こるとNa+が移動する(次図(ロ))。脱分極が続くと,活性化ゲー
トは開いているが,不活性化ゲートが閉じるためにイオンの移動は止まる(次図(ハ))。
再分極されると,不活性化ゲートが開き活性化ゲートが閉じて元の状態(次図(イ))
に戻る。
http://www.keirinkan.com/kori/kori_biology/kori_biology_n1/contents/bi-n1/4-bu/4-1-B.htm
活動電位(かつどうでんい、 action potential)は、なんらかの刺激に応じて
細胞膜に沿って流れる微弱な電位変化のこと。主としてナトリウムイオン、
カリウムイオンのイオンチャネルを通した受動的拡散でそれらイオンの細胞内外の
濃度差を変えることにより起きるものである。
細胞の内と外の間では、電位差が常に存在している。これは細胞内外でのイオン分布と、
これらイオンに対する細胞膜の透過性(特定のイオンの通しやすさ)に由来する。
電荷を持つイオンの分布が細胞内外で異なるため、活性化してない静止状態の
細胞の電位差は通常、細胞外と比べ細胞内がマイナスとなっている。
活動電位とは、この電位差がなんらかの刺激によって一時的に逆転する現象である。
活動電位はスパイクやインパルスともよばれる。また、活動電位に達することを
「発火する」(英 fire)と称することもある。
活動電位の速度と複雑さは細胞の種類により異なるものの、電位逆転の幅はだいたい同じである。
活動電位における負から正への電位の変化に要する時間は短く、数ミリ秒である。
どんな細胞の活動電位にも順に脱分極相、再分極相があり、多くの場合過分極相の段階もある。
活動電位は、太さ以外の条件が同じであるとき、より太い軸索上でより速く伝導する。
その速さは100-1000m/sである。
活動電位について知るためには、まず静止膜電位(静止電位ともいう)についておおまかに知る必要がある。
すべての細胞の膜内外に存在する電位差は通常、細胞外と比べ細胞内がマイナスである。
この状態を、膜は分極しているという。
活動していない状態にある膜の電位差は静止電位と呼ばれ、
神経細胞ではおよそ-70mVである(細胞外の電位を0としている)。
この電位差が生ずる要因はいくつかあるが、
もっとも重要なものは膜間における、イオン輸送と選択的なイオン透過性である。
活動電位の大まかな流れは次のようになっている。
[編集] 静止電位
静止電位において、いくつかのK+漏洩チャネルは開いている一方、
電位依存性Na+チャネルは閉じている。正味の電流は流れていないが、
膜間を移動している主なイオンはK+であり、その結果静止電位はK+平衡電位に比較近い値をとる。
[編集] 刺激
興奮刺激による膜の局所的な脱分極は、神経細胞の表面の膜にある電位依存性Na+チャネルを開く。
その結果Na+は濃度勾配および電気的勾配が推進力となり、細胞内へ流入する。
[編集] 脱分極
Na+が流入し膜電位の負電荷が減少するに従い、さらなるNa+チャネルが開き、
さらに大きなNa+の流入が引き起こされる。これは正のフィードバックのよい例である。
Na+チャネルが多く開くにつれ、Na+による電流はK+漏洩チャネルによる電流に打ち勝ち、
膜電位が逆転し内側がプラスとなる。
電位変化はNa+のみの移動によるものと考えるのではなく、
Na+とK+の膜間での「透過性の比」の変化によるものと考えるべきである。
この、脱分極とそれにともなうNa+チャネルの開口が周囲に広がっていくことで活動電位の伝導が起こる。
[編集] ピーク
膜電位が+30mV程度になると、Na+チャネルの電位感受性不活性化ゲートが閉じ、
さらなるNa+の流入を阻害する。それと同時に、
電位依存性K+チャネルの電位感受性活性化ゲートが開く。
活動電位のどの段階においても、きわめて少しのイオンの移動しか起こらないという
認識は重要である。活動電位が生じている間の、
細胞内外でのNa+とK+の濃度変化は無視できるほど極めて小さい。
http://ja.wikipedia.org/wiki/%E6%B4%BB%E5%8B%95%E9%9B%BB%E4%BD%8D
5、再分極
シナプス電位が+30mV程度になると、プラスイオンの流入が止まり、
逆にプラスイオンの流出が始まり急激にシナプス電位が元のマイナス電位に戻る
この再度マイナス電位に戻ることを再分極という
***参考資料***
再分極
電位依存性K+チャネルが開くことで、濃度勾配および電気的勾配が推進力となり
K+の流出が始まる。K+が流出することで、膜電位の逆転と再分極が引き起こされる。
http://ja.wikipedia.org/wiki/%E6%B4%BB%E5%8B%95%E9%9B%BB%E4%BD%8D
6、過分極
活動電位が立ち下がっているときはK+チャネルが完全に開いている状態なので、
K+の拡散電位:90mVにより、細胞内から細胞外へとK+イオン電流が流れる
K+の拡散電流は、
膜を通る電流 =
K+の90mVの拡散電位による拡散電流 + K+の膜電位による電圧電流=0
となる膜電位でとまることになり、
当然その膜電位は、-90mVとなる
このようにしてK+の平衡電位の-90mVになるまでシナプス電位が下がることになる
膜電位が通常の静止膜電位(-70mV)よりも低くなる状態を過分極という
再分極した後、一時的に過分極状態になる
他には、抑制性シナプス後電位によって過分極状態になることがある
***参考資料***
電位依存性K+チャネルの閉鎖は電位および時間に依存している。
このチャネルは膜電位の変化にすぐには応答せず、遅れて応答を返す。
そのため、膜が十分再分極した後もK+の流出が続き、一時的に膜電位が通常の静止電位より
もさらに低くなる。この過分極状態はアンダーシュートとも呼ばれる。
http://ja.wikipedia.org/wiki/%E6%B4%BB%E5%8B%95%E9%9B%BB%E4%BD%8D
7、不応期と活動電位の最大周波数
(1)絶対不応期と相対不応期
活動電位が発生した後、2ミリ秒程度は活動電位を発生できない。
この時期が絶対不応期という
(活動電位のパルスの立ち上がりから2ミリ秒であって、パルスの終了から2ミリ秒ではない
なので絶対不応期の2ミリ秒のうち1ミリ秒は活動電位のパルス波形の時間となる)
さらにその絶対不応期後の数ミリ秒は活動電位が発生しにくい。
この時期を相対不応期という
だいたい活動電位が発生した後の2ミリ秒から10ミリ秒ぐらい相対不応期がある
絶対不応期と相対不応期以上の期間では、
通常の大きな膜電位変化(約100mV)で活動電位を発生させることができるが、
相対不応期の活動電位の振幅は通常より小さくなる

不応期:インパルスが発生した後しばらくの間は、インパルスの発生できない期間を言う。
活動電位が発生してから2ミリ秒程度は活動電位発生できない。
この時期が絶対不応期(Absolute refractory period)。
その後数ミリ秒は活動電位が発生しにくい。
この時期が相対不応期(Relative refractory period)。
相対不応期には大きな膜電位変化で活動電位を発生させることができるが、
この時期の活動電位の振幅は通常より小さくなる。この図は神経線維(軸索)の束を
電気刺激(Stimulate)し、その神経線維上で離れた位置で記録(Record)したときの
様子を模式的に示している。左の図では、電気刺激の強さをS1からS4へと次第に強めている。
記録された電気活動は刺激強度が増すと次第に増加している。
右の図では2回の電気刺激(S1, S2)を行っている。S1とS2の時間間隔が小さくなると
応答の振幅が減少し、時間間隔が2ミリ秒以下のときは応答していない。
http://www.pri.kyoto-u.ac.jp/brain/brain/11/index-11.html
(2)活動電位の最大周波数
活動電位が発生できない絶対不応期期間
(活動電位の始まりから次の活動電位の始まりまでの期間)
:約2m秒
という情報から、活動電位の最大周波数は以下のようになる
活動電位の最大周波数:f=1/T=1/(2m秒)=約500Hz
(3)相対不応期における閾値電流変化
相対不応期の期間では、外部から活動電位を発生させるための
長方形閾値電流(電流:I=一定、時間幅:t=一定 の波形)の
閾値電流値(I)が変化しており
活動電位が発生したすぐ後(絶対不応期の約2ms後)は
かなり電流(I)がかなり高くないと活動電位が発生しない
図で表すと以下のようになる
活動電位が発生してから2ミリ秒程度は活動電位発生できない。
この時期が絶対不応期(Absolute refractory period)。
その後数ミリ秒は活動電位が発生しにくい。
この時期が相対不応期(Relative refractory period)。
相対不応期には大きな膜電位変化で活動電位を発生させることができるが、
この時期の活動電位の振幅は通常より小さくなる。この図は神経線維(軸索)の束を
電気刺激(Stimulate)し、その神経線維上で離れた位置で記録(Record)したときの
様子を模式的に示している。左の図では、電気刺激の強さをS1からS4へと次第に強めている。
記録された電気活動は刺激強度が増すと次第に増加している。
右の図では2回の電気刺激(S1, S2)を行っている。S1とS2の時間間隔が小さくなると
応答の振幅が減少し、時間間隔が2ミリ秒以下のときは応答していない。
http://www.pri.kyoto-u.ac.jp/brain/brain/11/index-11.html
(2)活動電位の最大周波数
活動電位が発生できない絶対不応期期間
(活動電位の始まりから次の活動電位の始まりまでの期間)
:約2m秒
という情報から、活動電位の最大周波数は以下のようになる
活動電位の最大周波数:f=1/T=1/(2m秒)=約500Hz
(3)相対不応期における閾値電流変化
相対不応期の期間では、外部から活動電位を発生させるための
長方形閾値電流(電流:I=一定、時間幅:t=一定 の波形)の
閾値電流値(I)が変化しており
活動電位が発生したすぐ後(絶対不応期の約2ms後)は
かなり電流(I)がかなり高くないと活動電位が発生しない
図で表すと以下のようになる
縦軸は、活動電位を発生させうるぎりぎりの刺激電流
横軸は、時間
強さ-期間曲線 strength-interval curve
この関係を式であらわすと、Weissの実験式がよくあてはまる。
この式は、刺激電流をI、刺激期間をtとすると、
I= a+ b/t
となる。この式において、t=∞ のときi=a。
それで、aは基電流を表す。
この式が相対不応期において活動電位を発生させるのに必要な電流となる
相対不応期で上式を考え直すと、tを
t:絶対不応期の終了時刻を0とした時間
(活動電位発生時刻からの時間:Tに直すなら t=T-2ms)
と考えると、相対不応期における
活動電位を発生させるのに必要な電流となる
このような特性となるのも、別の記事で等価回路と数式を用いて
解説します
(参照)
下記のサイトに
活動電位を発生させうるぎりぎりの刺激の強さを縦軸に、
時間を横軸にとった 強さ-期間曲線 strength-interval curve があり、
上式のように時間によって変化しているのがわかる
http://www.og-giken.co.jp/product/item/physical/cx-3.html
心筋活動電位には著名な 不応期 が認められ、絶対不応期 absolute r.p. と
相対不応期 relative r.p. とに分けられます。
活動電位を発生させ、種々の時期に種々の強さの刺激を与えて
もう1回活動電位を発生させうるかどうかを調べます。
活動電位を発生させうるぎりぎりの刺激の強さを縦軸に、
時間を横軸にとると 強さ-期間曲線 strength-interval curve が得られます。
この曲線から絶対および相対不応期を知ることができます。
http://www.mnc.toho-u.ac.jp/v-lab/shinkin/denki/denki-5-1.html
***参考資料***
Weissの実験式
http://www.tmd.ac.jp/med/phy1/ptext/stimulus.html
神経線維の電気信号発生のカギは、ニューロンをとりまく膜の内側と外側に存在する
イオンのアンバランスな分布にありました。細胞の内側には、カリウム・イオンが多く、
外側には、ナトリウム・イオンと塩素イオンが多く分布しています。
カリウムとナトリウムはプラスのイオンであり、塩素はマイナスのイオンでっす。
細胞の内側には、マイナスに帯電したタンパクが集まります。細胞が静止状態にあるとき、
膜の内側は、外側に対して、マイナス数十ミリボルト(1ミリボルトは、1000分の1ボルト)
の電位差を保たれます。これを、静止電位と呼んでいます。
このアンバランスなイオン分布を維持するためにニューロンは、エネルギーを使って
ナトリウムを細胞外に汲み出し、カリウムを細胞内に汲み入れる仕組みを持っています。
信号が、神経線維(神経細胞の情報を伝える突起=軸索)を通って伝えられるときには、
まず、細胞体の膜の電位が、ほんの一瞬、プラスに逆転します。
この膜の電位の逆転のきっかけは、他の神経細胞から信号を受け取った結果引き起こされる
こともあるし、また、外界からの物理的、
あるいは、化学的刺激によって引き起こされることもあります。
1000分の1秒以下の短い時間に引き起こされた、この膜の電位の逆転を
、 活動電位と呼んでいます。この一瞬の電位の逆転は、細胞の外からの急速なナトリウムの流入と、
それに続く細胞内からのカリウムの流出によって引き起こされたものです。
ほんの一瞬の膜の電位変化は、急速に起こり、速やかに元へ戻る。
オシロスコープでその電位変化を観察すると、鋭く尖った波形となるために、
インパルス、スパイク、発火、発射などと呼ばれています。
図1 活動電位:神経細胞の膜の内側は通常マイナス数十ミリ・ボルトですが、
膜の内側の電位がプラス方向にシフトしてその変化が一定のレベルに達すると、
まず、細胞外のナトリウム・イオンの細胞内への流入(緑の線)が起こり、
その直後に細胞内のカリウム・イオンの細胞外への流出(赤の線)が起こります。
この2つの変化が連続して起こることで、
細胞内の電位が1ミリ秒以下の短時間プラスになる現象が発生します。
これが活動電位(青の線)です。
細胞体にインパルス(活動電位)が発生するとき、膜の電位が次第に上昇し、
ある値(しきい値、閾値)を越えると、一気に発生します。インパルスの大きさは、
発生の時間間隔が極端に短くならない限りは、常に一定です。つまり、
発生するインパルスの大きさは、神経線維を遠くまで伝わっていくときにも
常に一定の大きさに保たれていて弱まることがない特徴を持っています。
これに対して、電線を電気が伝わるときは、信号が次第に弱まってしまいます。
そのために、電線を伝わる電気信号の場合は、
ところどころに信号を強めるための装置(増幅器)をつながなくてはなりません。
インパルスは、普通、細胞体から神経線維(軸索)がでていく付け根の部分でまず発生します。
この部分は、軸索がややふくらんで細胞体に連なっているので、軸索小丘と呼ばれています。
神経線維をインパルスが伝わっていくとき、発生したインパルスが近傍の神経線維の
膜の電位に電気的変化を引き起こします。この膜の電位の変化が決められた値(しきい値)
を越えることによってつぎつぎとインパルスが発生していきます。
インパルスがもと来た方へ逆戻りすることはありません。
インパルスが発生した後しばらくの間は、インパルスの発生できない不応期と
呼ばれる時期があるためです。こうして、神経線維を伝わるインパルスは常に順方向に、
通常は細胞体から次第に遠ざかる方向へと伝わっていきます。
http://www.pri.kyoto-u.ac.jp/brain/brain/11/index-11.html
8、活動電位から化学信号へ
活動電位が軸索の終末にくると、
シナプス前部には膜電位感受性カルシウムチャネルがありますので、
これが活動電位によって膜電位が上昇したことを感知して、活性化し、
つまりチャネルが開いてカルシウムが細胞内にはいります。
このカルシウムによって、シナプス前部にあったシナプス小胞が細胞膜に融合し、
中に入っていた神経伝達物質をシナプス間隙に放出します。
そして、別の神経細胞(ニューロン)にその神経伝達物質が伝達し、
シナプス電位を発生させます。
横軸は、時間
強さ-期間曲線 strength-interval curve
この関係を式であらわすと、Weissの実験式がよくあてはまる。
この式は、刺激電流をI、刺激期間をtとすると、
I= a+ b/t
となる。この式において、t=∞ のときi=a。
それで、aは基電流を表す。
この式が相対不応期において活動電位を発生させるのに必要な電流となる
相対不応期で上式を考え直すと、tを
t:絶対不応期の終了時刻を0とした時間
(活動電位発生時刻からの時間:Tに直すなら t=T-2ms)
と考えると、相対不応期における
活動電位を発生させるのに必要な電流となる
このような特性となるのも、別の記事で等価回路と数式を用いて
解説します
(参照)
下記のサイトに
活動電位を発生させうるぎりぎりの刺激の強さを縦軸に、
時間を横軸にとった 強さ-期間曲線 strength-interval curve があり、
上式のように時間によって変化しているのがわかる
http://www.og-giken.co.jp/product/item/physical/cx-3.html
心筋活動電位には著名な 不応期 が認められ、絶対不応期 absolute r.p. と
相対不応期 relative r.p. とに分けられます。
活動電位を発生させ、種々の時期に種々の強さの刺激を与えて
もう1回活動電位を発生させうるかどうかを調べます。
活動電位を発生させうるぎりぎりの刺激の強さを縦軸に、
時間を横軸にとると 強さ-期間曲線 strength-interval curve が得られます。
この曲線から絶対および相対不応期を知ることができます。
http://www.mnc.toho-u.ac.jp/v-lab/shinkin/denki/denki-5-1.html
***参考資料***
Weissの実験式
http://www.tmd.ac.jp/med/phy1/ptext/stimulus.html
神経線維の電気信号発生のカギは、ニューロンをとりまく膜の内側と外側に存在する
イオンのアンバランスな分布にありました。細胞の内側には、カリウム・イオンが多く、
外側には、ナトリウム・イオンと塩素イオンが多く分布しています。
カリウムとナトリウムはプラスのイオンであり、塩素はマイナスのイオンでっす。
細胞の内側には、マイナスに帯電したタンパクが集まります。細胞が静止状態にあるとき、
膜の内側は、外側に対して、マイナス数十ミリボルト(1ミリボルトは、1000分の1ボルト)
の電位差を保たれます。これを、静止電位と呼んでいます。
このアンバランスなイオン分布を維持するためにニューロンは、エネルギーを使って
ナトリウムを細胞外に汲み出し、カリウムを細胞内に汲み入れる仕組みを持っています。
信号が、神経線維(神経細胞の情報を伝える突起=軸索)を通って伝えられるときには、
まず、細胞体の膜の電位が、ほんの一瞬、プラスに逆転します。
この膜の電位の逆転のきっかけは、他の神経細胞から信号を受け取った結果引き起こされる
こともあるし、また、外界からの物理的、
あるいは、化学的刺激によって引き起こされることもあります。
1000分の1秒以下の短い時間に引き起こされた、この膜の電位の逆転を
、 活動電位と呼んでいます。この一瞬の電位の逆転は、細胞の外からの急速なナトリウムの流入と、
それに続く細胞内からのカリウムの流出によって引き起こされたものです。
ほんの一瞬の膜の電位変化は、急速に起こり、速やかに元へ戻る。
オシロスコープでその電位変化を観察すると、鋭く尖った波形となるために、
インパルス、スパイク、発火、発射などと呼ばれています。
図1 活動電位:神経細胞の膜の内側は通常マイナス数十ミリ・ボルトですが、
膜の内側の電位がプラス方向にシフトしてその変化が一定のレベルに達すると、
まず、細胞外のナトリウム・イオンの細胞内への流入(緑の線)が起こり、
その直後に細胞内のカリウム・イオンの細胞外への流出(赤の線)が起こります。
この2つの変化が連続して起こることで、
細胞内の電位が1ミリ秒以下の短時間プラスになる現象が発生します。
これが活動電位(青の線)です。
細胞体にインパルス(活動電位)が発生するとき、膜の電位が次第に上昇し、
ある値(しきい値、閾値)を越えると、一気に発生します。インパルスの大きさは、
発生の時間間隔が極端に短くならない限りは、常に一定です。つまり、
発生するインパルスの大きさは、神経線維を遠くまで伝わっていくときにも
常に一定の大きさに保たれていて弱まることがない特徴を持っています。
これに対して、電線を電気が伝わるときは、信号が次第に弱まってしまいます。
そのために、電線を伝わる電気信号の場合は、
ところどころに信号を強めるための装置(増幅器)をつながなくてはなりません。
インパルスは、普通、細胞体から神経線維(軸索)がでていく付け根の部分でまず発生します。
この部分は、軸索がややふくらんで細胞体に連なっているので、軸索小丘と呼ばれています。
神経線維をインパルスが伝わっていくとき、発生したインパルスが近傍の神経線維の
膜の電位に電気的変化を引き起こします。この膜の電位の変化が決められた値(しきい値)
を越えることによってつぎつぎとインパルスが発生していきます。
インパルスがもと来た方へ逆戻りすることはありません。
インパルスが発生した後しばらくの間は、インパルスの発生できない不応期と
呼ばれる時期があるためです。こうして、神経線維を伝わるインパルスは常に順方向に、
通常は細胞体から次第に遠ざかる方向へと伝わっていきます。
http://www.pri.kyoto-u.ac.jp/brain/brain/11/index-11.html
8、活動電位から化学信号へ
活動電位が軸索の終末にくると、
シナプス前部には膜電位感受性カルシウムチャネルがありますので、
これが活動電位によって膜電位が上昇したことを感知して、活性化し、
つまりチャネルが開いてカルシウムが細胞内にはいります。
このカルシウムによって、シナプス前部にあったシナプス小胞が細胞膜に融合し、
中に入っていた神経伝達物質をシナプス間隙に放出します。
そして、別の神経細胞(ニューロン)にその神経伝達物質が伝達し、
シナプス電位を発生させます。
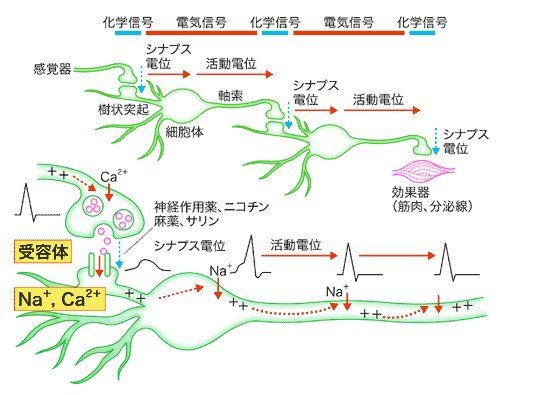
***参考資料***
活動電位からシナプス電位へ
シナプスにおいて、活動電位からシナプス電位に変換されます(図5)。
活動電位は、神経細胞の軸索突起の根元で発生し、軸索をつたわってシナプス前部に達します。
シナプス前部には膜電位感受性カルシウムチャネルがありますので、
これが活動電位によって膜電位が上昇したことを感知して、活性化し、
つまりチャネルが開いてカルシウムが細胞内にはいります。
このカルシウムによって、シナプス前部にあったシナプス小胞が細胞膜に融合し、
中に入っていた神経伝達物質をシナプス間隙に放出します。
興奮性の神経伝達を司っているグルタミン酸を例にあげてみますと、
グルタミン酸がシナプス後膜にあるその受容体、つまりグルタミン酸受容体に結合します。
グルタミン酸受容体は4つの膜蛋白からなり、それらのまん中に穴(チャネル)を形成していて、
受容体にグルタミン酸が結合するとタンパク質の構造が変化して、短時間穴が開きます。
この穴はカチオン(陽イオン;ナトリウム、カリウム、カルシウムなど)をとおす性質があるため、
細胞外には内側に比べて高濃度のナトリウムイオンがあり、
それがチャネルをとおって細胞内に流入します。
ナトリウムイオンはプラスですから、その結果、
細胞内のマイナスの程度が浅くなる、つまり脱分極します。
脱分極するにつれ、カリウムイオンの流出もあるために、
活動電位のように膜電位がプラスまで上昇することはありません。
このような神経伝達物質によって引き起されたシナプス後部の膜電位をシナプス電位といいます。
グルタミン酸 (Glu) の場合、シナプス電位は脱分極性なので興奮性シナプス後電位といいます。
一方、GABA (ギャバ) やグリシンは抑制性です。抑制性ニューロンからGABAが分泌されると、
GABA受容体チャネルが活性化し、塩素イオンを選択的に通す穴がひらき、
細胞外に多い塩素イオンが細胞内に流入します。塩素イオンはマイナスなので、
その結果膜電位はよりマイナスの方向に深くなり、つまり過分極になり、
これを抑制性シナプス後電位といいます。これによって、活動電位が出にくくなります(図5)。
麻酔薬、睡眠薬、抗不安薬などはこのGABA受容体の働きを促進させ、
グルタミン酸による興奮性の神経活動を抑制させるのです。
http://www.tmin.ac.jp/medical/17/synapse2.html
活動電位とシナプス電位
脳の活動は活動電位とシナプス電位によって成り立っています。
活動電位は細胞体からシナプス前部まで軸索突起(アクソン)を伝わります。
活動電位が軸索の終末にくるとシナプス前部から神経伝達物質が分泌され、
それが樹状突起(デンドライト)のシナプス後部の受容体を活性化し、
シナプス後細胞にシナプス電位を発生させます。
神経細胞は数千から数万ものシナプス入力をうけており、
それらのシナプス電位が統合され、軸索起部におけるその大きさによって
活動電位が発生するか否か、またその頻度がきまってきます。
つまり軸索部分の電気信号がシナプス部分で化学信号に変化し、
シナプス後部ニューロンでシナプス電位、さらに活動電位という電気信号に変化します。
このように神経系の情報伝達は電気信号(シナプス電位、活動電位)と
化学信号(神経伝達物質)の連鎖によります。
そしてシナプスとは神経伝達物質を介して、活動電位からシナプス電位に
変換する場であるのです。活動電位は軸索に存在するイオンチャネルの密度に規定され、
発生するか否かしかなく、一旦発生してしまえば大きさは一定です。
一方、シナプス電位はシナプ前部からの神経伝達物質の量やシナプス後部の
受容体の量的変化に依存して可変的です。
http://www.tmin.ac.jp/medical/17/synapse2.html
9、シナプスの可塑性
シナプスの活動状態などによってシナプスの伝達効率が変化するシナプス可塑性は、
記憶や学習に重要な役割を持つと考えられている。
シナプス前細胞とシナプス後細胞がともに高頻度で連続発火すると、
持続的なEPSPによりシナプスの伝達効率が増加する。
これを長期増強(LTP; Long Term Potentiation)という。
また、低頻度の発火や、抑制性シナプス後細胞の連続発火によるIPSPの持続によって、
シナプスの伝達効率が低下する現象を長期抑圧(LTD; Long Term Depression)という。
近年では、シナプス前細胞とシナプス後細胞の発火時間差のみによっても
結合強度に変化が見られることが分かっている。
これをスパイクタイミング依存シナプス可塑性
(STDP; Spike Timing Dependent Plasticity)という。
また、一旦LTPやLTDを起こしたシナプスに対して適切な刺激を与えると、
そのLTPやLTDが消失する事も知られており、
それぞれ脱増強 (Depotentiation)、脱抑圧(Dedepressiopn)などと呼ばれる。
http://ja.wikipedia.org/wiki/%E3%82%B7%E3%83%8A%E3%83%97%E3%82%B9
*** 神経細胞、活動電位、シナプス電位、化学信号などの参考サイト ***
膜電位
http://ja.wikipedia.org/wiki/%E8%86%9C%E9%9B%BB%E4%BD%8D
活動電位
http://ja.wikipedia.org/wiki/%E6%B4%BB%E5%8B%95%E9%9B%BB%E4%BD%8D
東京都医学研究機構 東京都神経科学総合研究所
http://www.tmin.ac.jp/medical/17/synapse2.html
ビジュアル生理学
http://bunseiri.hp.infoseek.co.jp/index.html
細胞生物学
http://www.tmd.ac.jp/artsci/biol/textbook/celltop.htm
BRAIN
http://kirigakure.dyndns.org/~brain/pukiwiki-1.4.5_1/index.php?BRAIN
脳の世界:京都大学霊長類研究所
http://www.pri.kyoto-u.ac.jp/brain/brain/index.html
神経生理テキスト
http://www.tmd.ac.jp/med/phy1/ptext/ptext_index.html
東京大学大学院理学系研究科 生体情報学研究室 岡 良隆教授の講義資料
http://www.biol.s.u-tokyo.ac.jp/users/naibunpi/Lecture_Index.html
活動電位からシナプス電位へ
シナプスにおいて、活動電位からシナプス電位に変換されます(図5)。
活動電位は、神経細胞の軸索突起の根元で発生し、軸索をつたわってシナプス前部に達します。
シナプス前部には膜電位感受性カルシウムチャネルがありますので、
これが活動電位によって膜電位が上昇したことを感知して、活性化し、
つまりチャネルが開いてカルシウムが細胞内にはいります。
このカルシウムによって、シナプス前部にあったシナプス小胞が細胞膜に融合し、
中に入っていた神経伝達物質をシナプス間隙に放出します。
興奮性の神経伝達を司っているグルタミン酸を例にあげてみますと、
グルタミン酸がシナプス後膜にあるその受容体、つまりグルタミン酸受容体に結合します。
グルタミン酸受容体は4つの膜蛋白からなり、それらのまん中に穴(チャネル)を形成していて、
受容体にグルタミン酸が結合するとタンパク質の構造が変化して、短時間穴が開きます。
この穴はカチオン(陽イオン;ナトリウム、カリウム、カルシウムなど)をとおす性質があるため、
細胞外には内側に比べて高濃度のナトリウムイオンがあり、
それがチャネルをとおって細胞内に流入します。
ナトリウムイオンはプラスですから、その結果、
細胞内のマイナスの程度が浅くなる、つまり脱分極します。
脱分極するにつれ、カリウムイオンの流出もあるために、
活動電位のように膜電位がプラスまで上昇することはありません。
このような神経伝達物質によって引き起されたシナプス後部の膜電位をシナプス電位といいます。
グルタミン酸 (Glu) の場合、シナプス電位は脱分極性なので興奮性シナプス後電位といいます。
一方、GABA (ギャバ) やグリシンは抑制性です。抑制性ニューロンからGABAが分泌されると、
GABA受容体チャネルが活性化し、塩素イオンを選択的に通す穴がひらき、
細胞外に多い塩素イオンが細胞内に流入します。塩素イオンはマイナスなので、
その結果膜電位はよりマイナスの方向に深くなり、つまり過分極になり、
これを抑制性シナプス後電位といいます。これによって、活動電位が出にくくなります(図5)。
麻酔薬、睡眠薬、抗不安薬などはこのGABA受容体の働きを促進させ、
グルタミン酸による興奮性の神経活動を抑制させるのです。
http://www.tmin.ac.jp/medical/17/synapse2.html
活動電位とシナプス電位
脳の活動は活動電位とシナプス電位によって成り立っています。
活動電位は細胞体からシナプス前部まで軸索突起(アクソン)を伝わります。
活動電位が軸索の終末にくるとシナプス前部から神経伝達物質が分泌され、
それが樹状突起(デンドライト)のシナプス後部の受容体を活性化し、
シナプス後細胞にシナプス電位を発生させます。
神経細胞は数千から数万ものシナプス入力をうけており、
それらのシナプス電位が統合され、軸索起部におけるその大きさによって
活動電位が発生するか否か、またその頻度がきまってきます。
つまり軸索部分の電気信号がシナプス部分で化学信号に変化し、
シナプス後部ニューロンでシナプス電位、さらに活動電位という電気信号に変化します。
このように神経系の情報伝達は電気信号(シナプス電位、活動電位)と
化学信号(神経伝達物質)の連鎖によります。
そしてシナプスとは神経伝達物質を介して、活動電位からシナプス電位に
変換する場であるのです。活動電位は軸索に存在するイオンチャネルの密度に規定され、
発生するか否かしかなく、一旦発生してしまえば大きさは一定です。
一方、シナプス電位はシナプ前部からの神経伝達物質の量やシナプス後部の
受容体の量的変化に依存して可変的です。
http://www.tmin.ac.jp/medical/17/synapse2.html
9、シナプスの可塑性
シナプスの活動状態などによってシナプスの伝達効率が変化するシナプス可塑性は、
記憶や学習に重要な役割を持つと考えられている。
シナプス前細胞とシナプス後細胞がともに高頻度で連続発火すると、
持続的なEPSPによりシナプスの伝達効率が増加する。
これを長期増強(LTP; Long Term Potentiation)という。
また、低頻度の発火や、抑制性シナプス後細胞の連続発火によるIPSPの持続によって、
シナプスの伝達効率が低下する現象を長期抑圧(LTD; Long Term Depression)という。
近年では、シナプス前細胞とシナプス後細胞の発火時間差のみによっても
結合強度に変化が見られることが分かっている。
これをスパイクタイミング依存シナプス可塑性
(STDP; Spike Timing Dependent Plasticity)という。
また、一旦LTPやLTDを起こしたシナプスに対して適切な刺激を与えると、
そのLTPやLTDが消失する事も知られており、
それぞれ脱増強 (Depotentiation)、脱抑圧(Dedepressiopn)などと呼ばれる。
http://ja.wikipedia.org/wiki/%E3%82%B7%E3%83%8A%E3%83%97%E3%82%B9
*** 神経細胞、活動電位、シナプス電位、化学信号などの参考サイト ***
膜電位
http://ja.wikipedia.org/wiki/%E8%86%9C%E9%9B%BB%E4%BD%8D
活動電位
http://ja.wikipedia.org/wiki/%E6%B4%BB%E5%8B%95%E9%9B%BB%E4%BD%8D
東京都医学研究機構 東京都神経科学総合研究所
http://www.tmin.ac.jp/medical/17/synapse2.html
ビジュアル生理学
http://bunseiri.hp.infoseek.co.jp/index.html
細胞生物学
http://www.tmd.ac.jp/artsci/biol/textbook/celltop.htm
BRAIN
http://kirigakure.dyndns.org/~brain/pukiwiki-1.4.5_1/index.php?BRAIN
脳の世界:京都大学霊長類研究所
http://www.pri.kyoto-u.ac.jp/brain/brain/index.html
神経生理テキスト
http://www.tmd.ac.jp/med/phy1/ptext/ptext_index.html
東京大学大学院理学系研究科 生体情報学研究室 岡 良隆教授の講義資料
http://www.biol.s.u-tokyo.ac.jp/users/naibunpi/Lecture_Index.html

コメント